
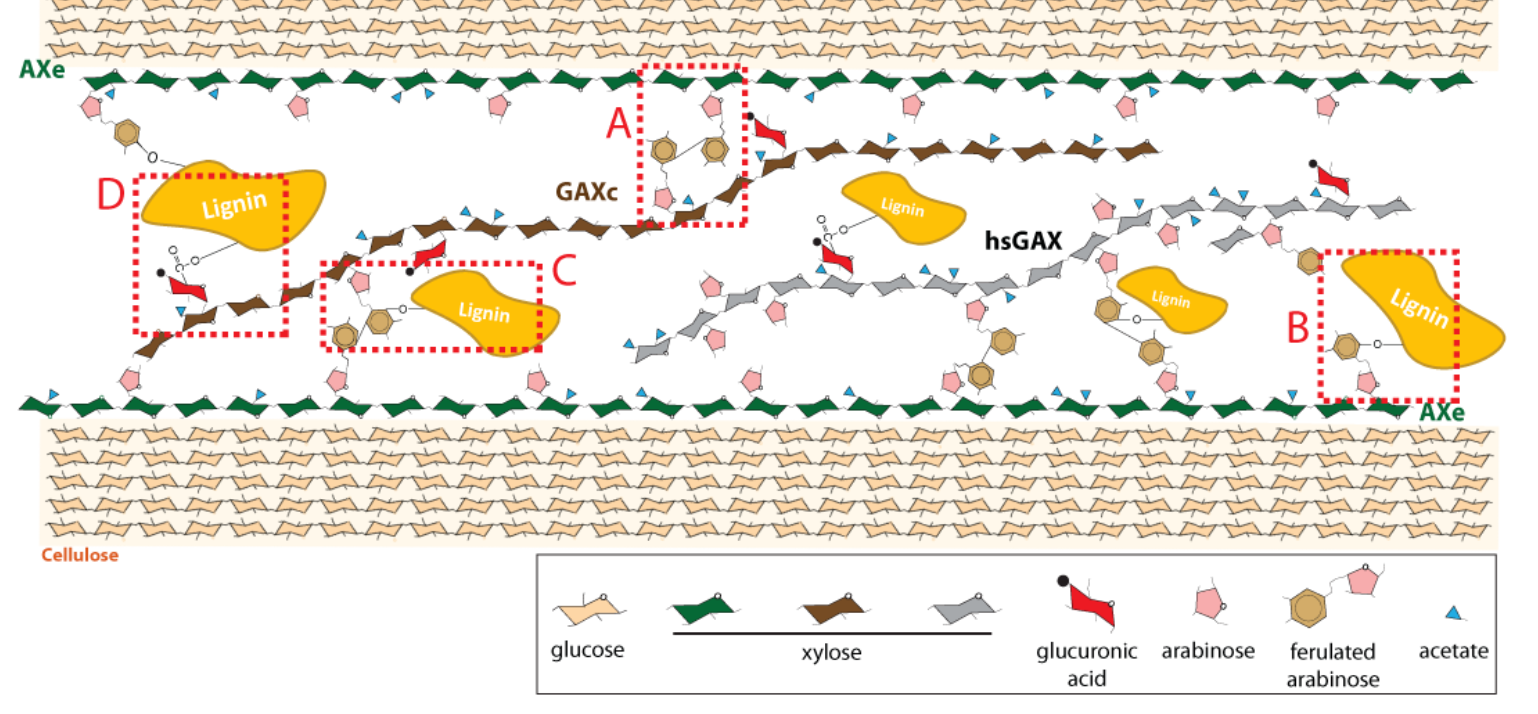
禾本科木聚糖中的一些阿拉伯呋喃糖是间隔均匀的,葡萄糖醛酸的修饰是成簇分布的。
在芒属茎秆木聚糖中,阿拉伯呋喃糖(Araf)的平均取代频率是11%,葡萄糖醛酸(GlcA)的替代是5%左右,但是这种取代是否随机,规则或无规则还不知道。
为了研究这些取代的距离,我们利用两种木聚糖酶(糖基水解酶(GH)家族GH5和GH30)水解了芒属茎秆的木聚糖.
GH5酶是阿拉伯木聚糖特异的木聚糖酶,主要是对应于阿拉伯呋喃糖侧链的水解。通过对GH5酶释放的寡糖进行定型,可以确定Araf取代之间的间隔.
GH30是葡萄糖醛酸酶,主要是对应葡萄糖醛酸的水解。
这些结果展示了两种十分不同的 禾本科木聚糖替换模式:a,阿拉伯木聚糖被阿拉伯呋喃糖均匀取代,b,葡萄糖醛酸阿拉伯木聚糖被葡萄糖醛酸成簇修饰。
这种模式在验证的8种禾本科作物是保守的。
禾本科木聚糖不同结构域的提取和分离
阿拉伯木聚糖和葡萄糖醛酸木聚糖都是细胞壁种的大量成分,在茎秆的细胞壁中,还发现一种可以被GH10水解的木聚糖,被称为高替换的葡萄糖醛酸木聚糖
为了了解木聚糖如何与其他成分互作,先是溶解消化提取多糖,然后PACE提取。
阿拉伯木聚糖的提取条件高于葡萄糖醛酸木聚糖,因此,作者认为阿拉伯木聚糖可能与细胞壁的结合更为紧密。
阿拉伯木聚糖和葡萄糖醛酸的带电性有差异,可能表明两者是存在不同的木聚糖分子中。
未被GH5和GH30水解的分子量小,hsGAX可能存在于小分子量的分子中。
阿拉伯木聚糖和葡萄糖醛酸的免疫共沉淀(二穗短柄草)
单克隆抗体可以连接形成荧光图像检测和定位细胞壁葡聚糖,高度敏感,通过特异识别寡糖结构。
结果表明这些细胞壁具有相对较高比例的频繁取代的木聚糖。
耐酶高取代木聚糖分布在细胞角落区域。
AXe和GAXc不局限于特定的细胞类型,存在于具有初级细胞壁和次级细胞壁的细胞中。

 浙公网安备 33010602011771号
浙公网安备 33010602011771号