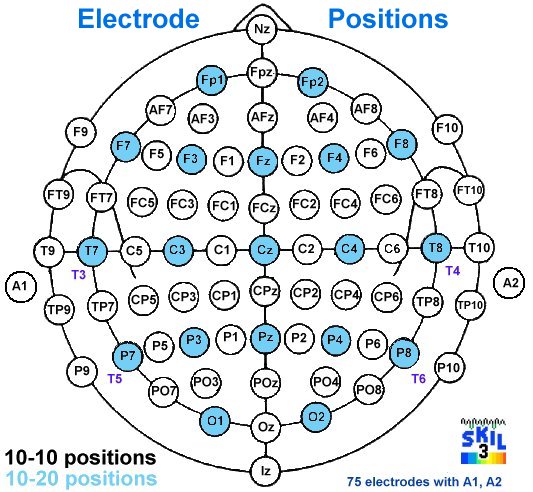
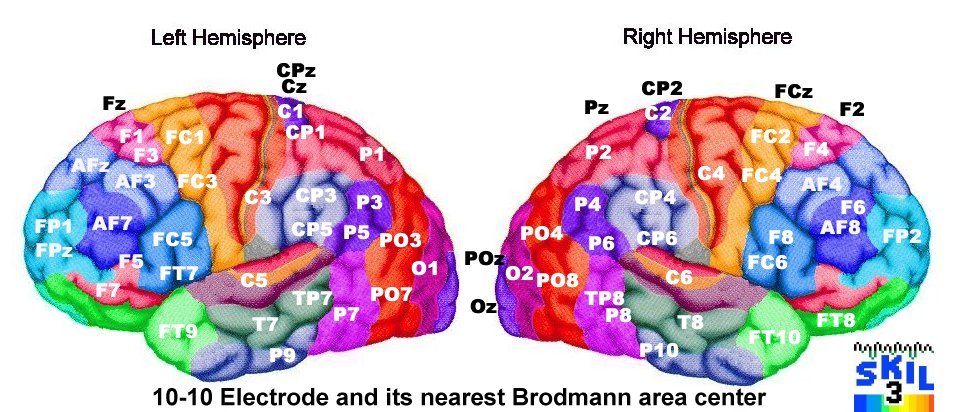
EEG: electrode positions & Broadmann atlas
Source: http://www.brainm.com/software/pubs/dg/BA_10-20_ROI_Talairach/nearesteeg.htm
|
|
* limited to SKIL Brodmann montage areas
- Closest Brodmann area to each 10-10 electrode
|
|
|
* limited to SKIL Brodmann montage areas
- The anatomical gyral names of the various Brodmann areas
Recall that there is a Brodmann area in both left and right hemisphere, the homologues.

Gyrus (Functional name)
1-3 – intermediate, caudal, and rostral postcentral (Primary Somatosensory Cortex)
4 - gigantopyramidal (Primary Motor Cortex)
5 - preparietal (Somatosensory Association Cortex)
6 - agranular frontal (Premotor cortex and Supplementary Motor Cortex)
7 - superior parietal (Somatosensory Association Cortex)
8 - intermediate frontal (includes Frontal eye fields)
9 - granular frontal (Dorsolateral prefrontal cortex, DLFC)
10 - frontopolar (DLFC)
11 - prefrontal (Orbitofrontal)
12 - prefrontal (Orbitofrontal)
17 - striate (Primary visual cortex, V1)
18 - parastriate (Secondary visual cortex, V2)
19 - peristriate (Tertiary or Associative visual cortex, V3)
20 - inferior temporal
21 - middle temporal
22 - superior temporal (caudal section considered Wernicke's area by most)
23 - ventral posterior cingulate
24 - ventral anterior cingulate
31 - dorsal posterior cingulate
32 - dorsal anterior cingulate
37 - occipitotemporal
38 - temporopolar (temporal pole)
39 – angular
40 - supramarginal
41-42 – ant. & posterior transverse temporal
44 - opercular (part of Broca's area on left hemisphere)
45 - triangular (part of Broca's area on left hemisphere)
46 - middle frontal
47 - orbital
- EXCLUDED FROM SKIL BRODMANN MONTAGE due to small size and distance from scalp
13 - insular
25 - subgenual
26 - ectosplenial
28 - entorhinal
29 - granular retrolimbic
30 - agranular retrolimbic
33 - pregenual
34 - dorsal entorhinal
35 - perirhinal
36 - ectorhinal
43 - subcentral
48 - retrosubicular
52 - parainsular
14, 15, 16, 27, 49, 50, 51 - monkey only Distance in mm. If you have a finding at an electrode, this table tells you which BA is closest
- 2nd table: If you have a BA you want to change, here is the electrode which is closest, distance in mm.
| FP1 | 10L,9L,46L,11L,32L |
|---|---|
| FPz | 10L,10R,9L,9R,11R |
| FP2 | 10R,9R,46R,11R,45R |
| AF7 | 46L,10L,45L,9L,11L |
| AF3 | 9L,46L,8L,10L,45L |
| AFz | 9L,9R,32R,32L,8L |
| AF4 | 9R,46R,8R,10R,45R |
| AF8 | 46R,10R,45R,9R,47R |
| F7 | 45L,47L,46L,44L,38L |
| F5 | 45L,46L,44L,47L,9L |
| F3 | 8L,6L,44L,45L,46L |
| F1 | 8L,6L,9L,32L,24L |
| Fz | 8L,8R,32L,24L,6R |
| F2 | 8R,6R,9R,32R,24R |
| F4 | 8R,9R,6R,44R,46R |
| F6 | 46R,44R,45R,9R,8R |
| F8 | 45R,47R,46R,44R,38R |
| FT9 | 38L,20L,21L,47L,22L |
| FT7 | 44L,47L,22L,38L,21L |
| FC5 | 44L,45L,22L,42L,41L |
| FC3 | 6L,4L,3L,44L,2L |
| FC1 | 6L,4L,5L,1L,2L |
| FCz | 6R,5L,6L,4L,5R |
| FC2 | 6R,4R,8R,2R,1R |
| FC4 | 6R,4R,44R,3R,2R |
| FC6 | 44R,45R,41R,22R,42R |
| FT8 | 47R,38R,44R,21R,22R |
| FT10 | 38R,20R,21R,47R,22R |
| T7 | 21L,42L,22L,41L,20L |
| C5 | 42L,41L,22L,40L,3L |
| C3 | 2L,4L,1L,3L,40L |
| C1 | 5L,2L,4L,1L,6L |
| Cz | 5L,5R,1R,1L,4L |
| C2 | 5R,1R,2R,4R,6R |
| C4 | 4R,2R,1R,3R,40R |
| C6 | 42R,41R,22R,3R,40R |
| T8 | 21R,22R,41R,42R,20R |
| TP7 | 21L,42L,22L,37L,41L |
| CP5 | 40L,39L,41L,42L,22L |
| CP3 | 40L,2L,1L,7L,3L |
| CP1 | 5L,7L,2L,1L,31L |
| CPz | 5L,5R,7R,7L,31L |
| CP2 | 5R,7R,1R,2R,40R |
| CP4 | 40R,1R,2R,5R,3R |
| CP6 | 40R,42R,39R,22R,41R |
| TP8 | 37R,21R,22R,42R,20R |
| P9 | 20L,37L,21L,22L,42L |
| P7 | 37L,39L,19L,21L,41L |
| P5 | 39L,19L,37L,40L,41L |
| P3 | 39L,7L,19L,40L,2L |
| P1 | 7L,31L,5L,7R,39L |
| Pz | 7R,7L,31R,31L,5R |
| P2 | 7R,31R,5R,19R,39R |
| P4 | 39R,40R,7R,19R,1R |
| P6 | 39R,19R,37R,40R,42R |
| P8 | 37R,39R,19R,42R,22R |
| P10 | 20R,37R,21R,22R,41R |
| PO7 | 19L,18L,37L,39L,17L |
| PO3 | 19L,39L,18L,17L,7L |
| POz | 17R,17L,18R,18L,7R |
| PO4 | 19R,39R,18R,17R,7R |
| PO8 | 19R,18R,39R,37R,17R |
| O1 | 18L,17L,19L,17R,39L |
| Oz | 17R,17L,18L,18R,19R |
| O2 | 18R,17R,19R,17L,39R |
| 1L | C3,C1,CP3,CP1,FC3 |
|---|---|
| 2L | C3,CP3,C1,CP1,FC3 |
| 3L | C3,C5,CP3,FC3,FC5 |
| 4L | C3,FC3,C1,FC1,CP3 |
| 5L | C1,CP1,CPz,Cz,C3 |
| 6L | FC3,FC1,F3,F1,C3 |
| 7L | P1,CP1,P3,Pz,CP3 |
| 8L | F1,F3,AF3,Fz,AFz |
| 9L | AF3,AFz,FP1,AF7,F3 |
| 10L | FP1,AF7,FPz,AF3,AFz |
| 11L | AF7,F7,FP1,FPz,AF3 |
| 17L | Oz,O1,POz,PO3,PO7 |
| 18L | O1,Oz,PO7,PO3,POz |
| 19L | PO7,PO3,P5,O1,P7 |
| 20L | FT9,T7,P9,TP7,FT7 |
| 21L | T7,TP7,FT7,C5,CP5 |
| 22L | T7,C5,TP7,FT7,CP5 |
| 23L | Pz,P1,CP1,POz,P3 |
| 24L | F1,Fz,F3,AFz,F2 |
| 31L | Pz,CP1,P1,CPz,CP3 |
| 32L | AFz,F1,Fz,AF3,F3 |
| 37L | P7,TP7,PO7,P5,P9 |
| 38L | FT9,FT7,F7,T7,F5 |
| 39L | P5,CP5,P3,P7,PO3 |
| 40L | CP3,CP5,C3,C5,P3 |
| 41L | C5,CP5,T7,TP7,FC5 |
| 42L | T7,C5,TP7,CP5,FT7 |
| 44L | FC5,F5,FT7,F7,F3 |
| 45L | F5,F7,AF7,FC5,FT7 |
| 46L | AF7,F5,F7,AF3,F3 |
| 47L | F7,FT7,AF7,F5,FT9 |
| 1R | C4,C2,CP4,CP2,FC2 |
| 2R | C4,C2,CP4,CP2,FC4 |
| 3R | C4,C6,CP4,FC4,CP6 |
| 4R | C4,FC4,C2,FC2,C6 |
| 5R | CP2,C2,CP4,CPz,C4 |
| 6R | FC2,FC4,F2,F4,C4 |
| 7R | P2,Pz,CP2,P4,CPz |
| 8R | F2,F4,AF4,Fz,F6 |
| 9R | AF4,AFz,F4,FP2,AF8 |
| 10R | FP2,AF8,FPz,AF4,F8 |
| 11R | FPz,FP2,AF8,F8,FP1 |
| 17R | Oz,O2,POz,PO4,PO8 |
| 18R | O2,Oz,PO8,PO4,POz |
| 19R | PO4,PO8,P6,O2,P4 |
| 20R | T8,FT10,TP8,P10,FT8 |
| 21R | T8,TP8,FT8,FT10,C6 |
| 22R | T8,C6,TP8,CP6,FT8 |
| 23R | Pz,P2,POz,CP2,P1 |
| 24R | F2,Fz,F4,AFz,F1 |
| 31R | Pz,P2,CP2,POz,CPz |
| 32R | AFz,F2,Fz,AF4,FPz |
| 37R | P8,TP8,PO8,P6,P10 |
| 38R | FT10,FT8,F8,T8,F6 |
| 39R | P6,PO4,P4,P8,PO8 |
| 40R | CP4,CP6,C4,P4,C6 |
| 41R | T8,C6,CP6,TP8,FT8 |
| 42R | C6,CP6,T8,TP8,FC6 |
| 44R | FC6,F6,F8,FC4,FT8 |
| 45R | F8,F6,AF8,FT8,AF4 |
| 46R | F6,AF8,F8,AF4,F4 |
| 47R | F8,FT8,AF8,FT10,F6 |
| FP1 | ba10L | 14 | ba09L | 28 | ba46L | 37 | ba11L | 38 | ba32L | 47 |
|---|---|---|---|---|---|---|---|---|---|---|
| FPz | ba10L | 25 | ba10R | 27 | ba09L | 34 | ba09R | 36 | ba11R | 38 |
| FP2 | ba10R | 16 | ba09R | 29 | ba46R | 35 | ba11R | 39 | ba45R | 43 |
| AF7 | ba46L | 16 | ba10L | 22 | ba45L | 28 | ba09L | 29 | ba11L | 32 |
| AF3 | ba09L | 15 | ba46L | 23 | ba08L | 25 | ba10L | 30 | ba45L | 32 |
| AFz | ba09L | 26 | ba09R | 27 | ba32R | 33 | ba32L | 33 | ba08L | 34 |
| AF4 | ba09R | 14 | ba46R | 21 | ba08R | 27 | ba10R | 29 | ba45R | 32 |
| AF8 | ba46R | 18 | ba10R | 20 | ba45R | 26 | ba09R | 29 | ba47R | 35 |
| F7 | ba45L | 13 | ba47L | 15 | ba46L | 17 | ba44L | 27 | ba38L | 32 |
| F5 | ba45L | 13 | ba46L | 17 | ba44L | 19 | ba47L | 33 | ba09L | 36 |
| F3 | ba08L | 21 | ba06L | 26 | ba44L | 30 | ba45L | 31 | ba46L | 32 |
| F1 | ba08L | 13 | ba06L | 28 | ba09L | 32 | ba32L | 36 | ba24L | 36 |
| Fz | ba08L | 27 | ba08R | 32 | ba32L | 37 | ba24L | 38 | ba06R | 38 |
| F2 | ba08R | 16 | ba06R | 27 | ba09R | 31 | ba32R | 38 | ba24R | 39 |
| F4 | ba08R | 18 | ba09R | 28 | ba06R | 28 | ba44R | 28 | ba46R | 28 |
| F6 | ba46R | 16 | ba44R | 19 | ba45R | 19 | ba09R | 32 | ba08R | 32 |
| F8 | ba45R | 14 | ba47R | 16 | ba46R | 19 | ba44R | 27 | ba38R | 30 |
| FT9 | ba38L | 22 | ba20L | 26 | ba21L | 35 | ba47L | 37 | ba22L | 41 |
| FT7 | ba44L | 26 | ba47L | 29 | ba22L | 29 | ba38L | 29 | ba21L | 30 |
| FC5 | ba44L | 13 | ba45L | 29 | ba22L | 34 | ba42L | 34 | ba41L | 34 |
| FC3 | ba06L | 19 | ba04L | 21 | ba03L | 31 | ba44L | 31 | ba02L | 34 |
| FC1 | ba06L | 24 | ba04L | 24 | ba05L | 34 | ba01L | 35 | ba02L | 36 |
| FCz | ba06R | 37 | ba05L | 41 | ba06L | 42 | ba04L | 44 | ba05R | 44 |
| FC2 | ba06R | 20 | ba04R | 29 | ba08R | 36 | ba02R | 36 | ba01R | 37 |
| FC4 | ba06R | 21 | ba04R | 21 | ba44R | 28 | ba03R | 33 | ba02R | 34 |
| FC6 | ba44R | 12 | ba45R | 33 | ba41R | 33 | ba22R | 34 | ba42R | 34 |
| FT8 | ba47R | 25 | ba38R | 27 | ba44R | 28 | ba21R | 28 | ba22R | 31 |
| FT10 | ba38R | 24 | ba20R | 27 | ba21R | 30 | ba47R | 36 | ba22R | 43 |
| T7 | ba21L | 14 | ba42L | 15 | ba22L | 16 | ba41L | 25 | ba20L | 27 |
| C5 | ba42L | 19 | ba41L | 20 | ba22L | 22 | ba40L | 25 | ba03L | 27 |
| C3 | ba02L | 16 | ba04L | 17 | ba01L | 20 | ba03L | 21 | ba40L | 25 |
| C1 | ba05L | 16 | ba02L | 23 | ba04L | 23 | ba01L | 23 | ba06L | 37 |
| Cz | ba05L | 29 | ba05R | 32 | ba01R | 43 | ba01L | 44 | ba04L | 45 |
| C2 | ba05R | 18 | ba01R | 23 | ba02R | 25 | ba04R | 28 | ba06R | 34 |
| C4 | ba04R | 17 | ba02R | 19 | ba01R | 22 | ba03R | 22 | ba40R | 26 |
| C6 | ba42R | 19 | ba41R | 22 | ba22R | 23 | ba03R | 28 | ba40R | 29 |
| T8 | ba21R | 13 | ba22R | 18 | ba41R | 21 | ba42R | 23 | ba20R | 26 |
| TP7 | ba21L | 20 | ba42L | 23 | ba22L | 25 | ba37L | 25 | ba41L | 28 |
| CP5 | ba40L | 20 | ba39L | 21 | ba41L | 23 | ba42L | 25 | ba22L | 29 |
| CP3 | ba40L | 19 | ba02L | 20 | ba01L | 24 | ba07L | 28 | ba03L | 30 |
| CP1 | ba05L | 19 | ba07L | 20 | ba02L | 26 | ba01L | 28 | ba31L | 37 |
| CPz | ba05L | 29 | ba05R | 30 | ba07R | 31 | ba07L | 34 | ba31L | 39 |
| CP2 | ba05R | 15 | ba07R | 23 | ba01R | 26 | ba02R | 30 | ba40R | 38 |
| CP4 | ba40R | 15 | ba01R | 25 | ba02R | 25 | ba05R | 29 | ba03R | 30 |
| CP6 | ba40R | 20 | ba42R | 22 | ba39R | 28 | ba22R | 28 | ba41R | 29 |
| TP8 | ba37R | 22 | ba21R | 24 | ba22R | 27 | ba42R | 27 | ba20R | 28 |
| P9 | ba20L | 29 | ba37L | 35 | ba21L | 42 | ba22L | 52 | ba42L | 53 |
| P7 | ba37L | 17 | ba39L | 24 | ba19L | 27 | ba21L | 39 | ba41L | 40 |
| P5 | ba39L | 9 | ba19L | 21 | ba37L | 29 | ba40L | 34 | ba41L | 40 |
| P3 | ba39L | 23 | ba07L | 24 | ba19L | 31 | ba40L | 33 | ba02L | 39 |
| P1 | ba07L | 15 | ba31L | 38 | ba05L | 40 | ba07R | 41 | ba39L | 41 |
| Pz | ba07R | 21 | ba07L | 26 | ba31R | 33 | ba31L | 36 | ba05R | 41 |
| P2 | ba07R | 15 | ba31R | 35 | ba05R | 36 | ba19R | 38 | ba39R | 39 |
| P4 | ba39R | 22 | ba40R | 28 | ba07R | 29 | ba19R | 30 | ba01R | 39 |
| P6 | ba39R | 10 | ba19R | 24 | ba37R | 29 | ba40R | 32 | ba42R | 39 |
| P8 | ba37R | 16 | ba39R | 24 | ba19R | 31 | ba42R | 40 | ba22R | 42 |
| P10 | ba20R | 32 | ba37R | 32 | ba21R | 42 | ba22R | 54 | ba41R | 56 |
| PO7 | ba19L | 18 | ba18L | 25 | ba37L | 26 | ba39L | 29 | ba17L | 35 |
| PO3 | ba19L | 19 | ba39L | 28 | ba18L | 28 | ba17L | 31 | ba07L | 37 |
| POz | ba17R | 30 | ba17L | 30 | ba18R | 36 | ba18L | 36 | ba07R | 37 |
| PO4 | ba19R | 15 | ba39R | 20 | ba18R | 28 | ba17R | 33 | ba07R | 37 |
| PO8 | ba19R | 22 | ba18R | 24 | ba39R | 26 | ba37R | 27 | ba17R | 34 |
| O1 | ba18L | 12 | ba17L | 15 | ba19L | 23 | ba17R | 37 | ba39L | 41 |
| Oz | ba17R | 12 | ba17L | 13 | ba18L | 24 | ba18R | 24 | ba19R | 40 |
| O2 | ba18R | 12 | ba17R | 15 | ba19R | 26 | ba17L | 36 | ba39R | 38 |
| ba01L | C3 | 20 | C1 | 23 | CP3 | 24 | CP1 | 28 | FC3 | 34 |
|---|---|---|---|---|---|---|---|---|---|---|
| ba02L | C3 | 16 | CP3 | 20 | C1 | 23 | CP1 | 26 | FC3 | 34 |
| ba03L | C3 | 21 | C5 | 27 | CP3 | 30 | FC3 | 31 | FC5 | 34 |
| ba04L | C3 | 17 | FC3 | 21 | C1 | 23 | FC1 | 24 | CP3 | 36 |
| ba05L | C1 | 16 | CP1 | 19 | CPz | 29 | Cz | 29 | C3 | 32 |
| ba06L | FC3 | 19 | FC1 | 24 | F3 | 26 | F1 | 28 | C3 | 32 |
| ba07L | P1 | 15 | CP1 | 20 | P3 | 24 | Pz | 26 | CP3 | 28 |
| ba08L | F1 | 13 | F3 | 21 | AF3 | 25 | Fz | 27 | AFz | 34 |
| ba09L | AF3 | 15 | AFz | 26 | FP1 | 28 | AF7 | 29 | F3 | 32 |
| ba10L | FP1 | 14 | AF7 | 22 | FPz | 25 | AF3 | 30 | AFz | 39 |
| ba11L | AF7 | 32 | F7 | 34 | FP1 | 38 | FPz | 46 | AF3 | 49 |
| ba17L | Oz | 13 | O1 | 15 | POz | 30 | PO3 | 31 | PO7 | 35 |
| ba18L | O1 | 12 | Oz | 24 | PO7 | 25 | PO3 | 28 | POz | 36 |
| ba19L | PO7 | 18 | PO3 | 19 | P5 | 21 | O1 | 23 | P7 | 27 |
| ba20L | FT9 | 26 | T7 | 27 | P9 | 29 | TP7 | 30 | FT7 | 37 |
| ba21L | T7 | 14 | TP7 | 20 | FT7 | 30 | C5 | 32 | CP5 | 34 |
| ba22L | T7 | 16 | C5 | 22 | TP7 | 25 | FT7 | 29 | CP5 | 29 |
| ba23L | Pz | 43 | P1 | 45 | CP1 | 47 | POz | 47 | P3 | 49 |
| ba24L | F1 | 36 | Fz | 38 | F3 | 41 | AFz | 44 | F2 | 44 |
| ba31L | Pz | 36 | CP1 | 37 | P1 | 38 | CPz | 39 | CP3 | 44 |
| ba32L | AFz | 33 | F1 | 36 | Fz | 37 | AF3 | 38 | F3 | 41 |
| ba37L | P7 | 17 | TP7 | 25 | PO7 | 26 | P5 | 29 | P9 | 35 |
| ba38L | FT9 | 22 | FT7 | 29 | F7 | 32 | T7 | 45 | F5 | 50 |
| ba39L | P5 | 9 | CP5 | 21 | P3 | 23 | P7 | 24 | PO3 | 28 |
| ba40L | CP3 | 19 | CP5 | 20 | C3 | 25 | C5 | 25 | P3 | 33 |
| ba41L | C5 | 20 | CP5 | 23 | T7 | 25 | TP7 | 28 | FC5 | 34 |
| ba42L | T7 | 15 | C5 | 19 | TP7 | 23 | CP5 | 25 | FT7 | 31 |
| ba44L | FC5 | 13 | F5 | 19 | FT7 | 26 | F7 | 27 | F3 | 30 |
| ba45L | F5 | 13 | F7 | 13 | AF7 | 28 | FC5 | 29 | FT7 | 31 |
| ba46L | AF7 | 16 | F5 | 17 | F7 | 17 | AF3 | 23 | F3 | 32 |
| ba47L | F7 | 15 | FT7 | 29 | AF7 | 32 | F5 | 33 | FT9 | 37 |
| ba01R | C4 | 22 | C2 | 23 | CP4 | 25 | CP2 | 26 | FC2 | 37 |
| ba02R | C4 | 19 | C2 | 25 | CP4 | 25 | CP2 | 30 | FC4 | 34 |
| ba03R | C4 | 22 | C6 | 28 | CP4 | 30 | FC4 | 33 | CP6 | 34 |
| ba04R | C4 | 17 | FC4 | 21 | C2 | 28 | FC2 | 29 | C6 | 35 |
| ba05R | CP2 | 15 | C2 | 18 | CP4 | 29 | CPz | 30 | C4 | 32 |
| ba06R | FC2 | 20 | FC4 | 21 | F2 | 27 | F4 | 28 | C4 | 33 |
| ba07R | P2 | 15 | Pz | 21 | CP2 | 23 | P4 | 29 | CPz | 31 |
| ba08R | F2 | 16 | F4 | 18 | AF4 | 27 | Fz | 32 | F6 | 32 |
| ba09R | AF4 | 14 | AFz | 27 | F4 | 28 | FP2 | 29 | AF8 | 29 |
| ba10R | FP2 | 16 | AF8 | 20 | FPz | 27 | AF4 | 29 | F8 | 38 |
| ba11R | FPz | 38 | FP2 | 39 | AF8 | 41 | F8 | 46 | FP1 | 51 |
| ba17R | Oz | 12 | O2 | 15 | POz | 30 | PO4 | 33 | PO8 | 34 |
| ba18R | O2 | 12 | Oz | 24 | PO8 | 24 | PO4 | 28 | POz | 36 |
| ba19R | PO4 | 15 | PO8 | 22 | P6 | 24 | O2 | 26 | P4 | 30 |
| ba20R | T8 | 26 | FT10 | 27 | TP8 | 28 | P10 | 32 | FT8 | 38 |
| ba21R | T8 | 13 | TP8 | 24 | FT8 | 28 | FT10 | 30 | C6 | 36 |
| ba22R | T8 | 18 | C6 | 23 | TP8 | 27 | CP6 | 28 | FT8 | 31 |
| ba23R | Pz | 45 | P2 | 47 | POz | 47 | CP2 | 49 | P1 | 50 |
| ba24R | F2 | 39 | Fz | 39 | F4 | 43 | AFz | 43 | F1 | 44 |
| ba31R | Pz | 33 | P2 | 35 | CP2 | 39 | POz | 40 | CPz | 41 |
| ba32R | AFz | 33 | F2 | 38 | Fz | 39 | AF4 | 40 | FPz | 41 |
| ba37R | P8 | 16 | TP8 | 22 | PO8 | 27 | P6 | 29 | P10 | 32 |
| ba38R | FT10 | 24 | FT8 | 27 | F8 | 30 | T8 | 44 | F6 | 49 |
| ba39R | P6 | 10 | PO4 | 20 | P4 | 22 | P8 | 24 | PO8 | 26 |
| ba40R | CP4 | 15 | CP6 | 20 | C4 | 26 | P4 | 28 | C6 | 29 |
| ba41R | T8 | 21 | C6 | 22 | CP6 | 29 | TP8 | 30 | FT8 | 31 |
| ba42R | C6 | 19 | CP6 | 22 | T8 | 23 | TP8 | 27 | FC6 | 34 |
| ba44R | FC6 | 12 | F6 | 19 | F8 | 27 | FC4 | 28 | FT8 | 28 |
| ba45R | F8 | 14 | F6 | 19 | AF8 | 26 | FT8 | 32 | AF4 | 32 |
| ba46R | F6 | 16 | AF8 | 18 | F8 | 19 | AF4 | 21 | F4 | 28 |
| ba47R | F8 | 16 | FT8 | 25 | AF8 | 35 | FT10 | 36 | F6 | 36 |
left ba01=right thumb activity (WOEXP: 497)!!
left ba02= Active right middle finger movement versus rest (WOEXP: 268)
left ba03=imitating symbolic finger movements (WOEXP: 145)
left ba04=
left ba05=
left ba06=
left ba07=
left ba08= Judge basic elements of reading ( Lower/upper case v. syllable WOEXP: 553)
left ba09=
left ba10=
left ba11=
left ba17= Visual exploration versus saccades (WOEXP: 6)
left ba18= Verbal numerical notation (WOEXP: 23)
left ba19=Visual motion (WOEXP: 430)!
left ba20=Visual categorization (WOEXP: 4)!!
left ba21=Visual meaningfulness (WOEXP: 164)
left ba22=
left ba23=Answering self-reflective v. semantic questions -w/anterior site (WOEXP: 62)
left ba24=part of pain sensitivity network (WOEXP: 238)
left ba31=
left ba32=Early phase heat pain (WOEXP: 298)
left ba37=
left ba38=
left ba39=Motion verb sentences versus static sentences (WOEXP: 534) !
left ba40=
left ba41=
left ba42=
left ba44=
left ba45=Word generation (WOEXP: 32)
left ba46=
left ba47=
right ba01=imperceptible electric finger stimulation (WOEXP: 278)
right ba02=
right ba03=
right ba04=
right ba05=
right ba06=
right ba07=Mental rotation of figures versus object determination or dots counting (WOEXP: 86)
right ba08= Sensorimotor willed action (WOEXP: 13)
right ba09=
right ba10=
right ba11=
right ba17=Large line patterns (WOEXP: 101) !!
right ba18=Verbal numerical notation (WOEXP: 23) !
right ba19=
right ba20=Negative interaction between predictable tones and button press (WOEXP: 261) !!
right ba21=Spatial neglect (WOEXP: 185) !!
right ba22=
right ba23=Task-related episodic retrieval versus semantic (WOEXP: 565)
right ba24=
right ba31=Correlation with pain intensity (WOEXP: 248) !! Decreases in heat pain in left forearm (WOEXP: 363)
right ba32=
right ba37=
right ba38=
right ba39=Biological visual motion (WOEXP: 111) !
right ba40=
right ba41=Auditory change (WOEXP: 454) ! Chords (WOEXP: 22) !!
right ba42=
right ba44= sole right side of network involved in Response competition (WOEXP: 134)
right ba45=
right ba46= Decreases during 100 Hz vibration on left forearm (WOEXP: 365)
right ba47=
=====
Lloyd table 1.3 >8% normalized only, EEG accessible
L 3 13.3 Action.Motor Learning
L 4 11.4 Action.Motor Learning
L 6 8.6 Cognition.Time
L 7 8.3 Action.Motor Learning
L 9 8.0 Cognition.Time
L 17 12.0 Perception.Vision.Color
L 21 12.0 Action.Motor Learning
L 37 9.6 Emotion.Anxiety
L 47 8.9 Cognition.Time
R 1 8.2 Perception.Audition
R 6 8.2 Action.Observation
R 9 9.6 Action.Motor Learning
R 17 8.0 Perception.Vision.Color
R 22 8.0 Perception.Olfaction
R 24 13.3 Action.Motor Learning
R 37 8.6 Perception.Vision.Color
R 38 9.4 Interoception.Sexuality
R 40 9.2 Action.Motor Learning
R 43 17.1 Interoception.Hunger
R 47 9.6 Perception.Olfaction
cytoarchitecture and neurotransmitter-binding site distributions divide BA 4 into anterior and posterior sites
voluntary movements differently modulate the somatosensory functions of SMA and SM1 (Mima et al., 1999)
=======================================
- Brodmann's Interactive AtlasFunction: Brodmann's Areas
.. Motor
Primary motor: 4, 1, 2, 3
Secondary motor: 6, 8
Motor planning: 6, 13-16; 24, 32-33; 40
Motor Imagery: 5, 7, 4, 6, 8; 24, 32-33
Motor Learning: 4, 1-3, 6, 8; 23, 26, 29-31
Saccadic movements: 4, 5, 7, 6, 8, 17, 18, 19, 46
Inhibition of blinking: 4
.. Sensory
Proprioception: 1-3, 4, 8
Touch, temperature, vibration: 1-3, 4, 5, 7, 13-16
Somatosensory integration: 40
.. Auditory
Basic processing: 41, 42
Complex sounds processing: 21, 22
Auditory Imagery: 8, 9, 10
Familiar voices: 38
.. Visual
Light intensity / patterns: 17, 18, 19
Color discrimination: 17
Visual integration: 20
Visual motion processing: 37
.. Olfaction
General olfaction: 11
Familiar odors: 9, 10; 24, 32-33; 44, 45, 47
.. Language
Comprehension: 22, 20, 21, 37, 39, 40, 5, 7, 6, 9, 10, 23, 26, 29-31, 38, 43, 44, 45,47
Expression: 44, 45, 46, 6, 8, 9, 10, 13-16, 21; 24, 32-33; 47
Prosody comprehension: 22
Reading: 6, 39
Writting: 40
.. Memory
Working Memory: 5, 7, 6, 8, 9, 10, 20; 24, 32-33; 40, 41, 44, 45, 46, 47; (27-28, 34-36, 48)
Episodic memory: 6, 44, 45, 47
Retrieval: 8, 9, 10,; 26, 29, 29-31; 24, 32-33; 38, 40
Encoding: (27-28, 34-36, 48); 9, 10; 24, 32-33; 37, 46
Topokinetic: 23, 26, 29-31
.. Attention
Visual: 17, 18, 37
Visuomotor: 5, 7, 6, 8
Visuospatial: 6, 8; 39, 24, 32-33; 45
Selective to sounds: 6, 9, 10,; 24, 32-33
To speech: 20, 22,; 23, 26, 29-31; 38, 47
.. Executive
Planning: 6, 8, 9, 10
Behavioral inhibition: 6, 8, 9, 10, 13-16; 24, 32-33; 39, 40, 44 , 46, 47
Motor inhibition: 24, 32-33, 44, 45, 47
.. Emotion
Experiencing / processing emotion: 38, 46; (27-28, 34-36, 48)
Related to language: 23, 26, 29-31; 25
Emotional stimuli: 9, 10; 24, 32-33
Fear response: 13-16
..Pain
Pain processing: 13-16; 24, 32-33, 5, 7
.. Others
Calculation: 39, 40, 6, 8, 9, 10, 13-16, 46
Theory of mind: 38, 9, 10, 20, 21, 22, 37, 47
Face recognition: 37
Mental time-keeping: 24, 32-33
Sexual arousal: 24, 32-33
Humor comprehension: 38
Music performance: 40
Music enjoyment: 44, 45, 46
Navegational skills: (27-28, 34-36, 48)
Novelty discrimination: (27-28, 34-36, 48)
==========================================
---WOEXP: 4. ----
Neuropsychologia. 2000;38(13):1693-703.
Categorization and category effects in normal object recognition: a PET study.
Gerlach C, Law I, Gade A, Paulson OB.
To investigate the neural correlates of the structural and semantic stages of visual object recognition and to see whether any effects of category could be found at these stages, we compared the rCBF associated with two categorization tasks (subjects decided whether pictures represented artefacts or natural objects), and two object decision tasks (subjects decided whether pictures represented real objects or nonobjects). The categorization tasks differed from each other in that the items presented in the critical scan window were drawn primarily from the category of artefacts in the one task and from the category of natural objects in the other. The same was true for the object decision tasks. The experiment thus comprised a two-by-two factorial design. The factors were Task Type with two levels (object decision vs. categorization) and Category also with two levels (natural objects vs. artefacts). The object decision tasks were associated with activation of areas involved in structural processing (fusiform gyri, right inferior frontal gyrus). In contrast, the categorization tasks were associated with activation of the left inferior temporal gyrus, a structure believed to be involved in semantic processing. In addition, activation of the left premotor cortex was found during the categorization of artefacts compared with both the categorization of natural objects and object decision to artefacts. These findings suggest that the structural and semantic stages are dissociable and that the categorization of artefacts, as opposed to the categorization of natural objects, is based, in part, on action knowledge mediated by the left premotor cortex. However, because artefacts and natural objects often caused activation in the same regions within tasks, processing of these categories is not totally segregated. Rather, the categories differ in their weight on different forms of knowledge in particular tasks.
---164---
Brain. 1997 Oct;120 ( Pt 10):1763-77.
Brain activity during observation of actions. Influence of action content and subject's strategy.
Decety J, Grèzes J, Costes N, Perani D, Jeannerod M, Procyk E, Grassi F, Fazio F.
Processus mentaux et activation cérébrale, Inserm Unit, Bron, France.
PET was used to map brain regions that are associated with the observation of meaningful and meaningless hand actions. Subjects were scanned under four conditions which consisted of visually presented actions. In each of the four experimental conditions, they were instructed to watch the actions with one of two aims: to be able to recognize or to imitate them later. We found that differences in the meaning of the action, irrespective of the strategy used during observation, lead to different patterns of brain activity and clear left/right asymmetries. Meaningful actions strongly engaged the left hemisphere in frontal and temporal regions while meaningless actions involved mainly the right occipitoparietal pathway. Observing with the intent to recognize activated memory-encoding structures. In contrast, observation with the intent to imitate was associated with activation in the regions involved in the planning and in the generation of actions. Thus, the pattern of brain activation during observation of actions is dependent both on the nature of the required executive processing and the type of the extrinsic properties of the action presented.
Observation of meaningful action in order to recognize versus observation of meaningless action. Observation of hand and arm meaningful action such as "opening a bottle", "drawing a line", "sewing a button" showed on a video for later recognition. WOEXP: 164.
---62---
Brain. 2002 Aug;125(Pt 8):1808-14.
Neural correlates of self-reflection.
Johnson SC, Baxter LC, Wilder LS, Pipe JG, Heiserman JE, Prigatano GP.
Department of Clinical Neuropsychology, Barrow Neurological Institute, St Joseph's Hospital and Medical Center, Phoenix, AZ 85013, USA. s2johns@chw.edu
The capacity to reflect on one's sense of self is an important component of self-awareness. In this paper, we investigate some of the neurocognitive processes underlying reflection on the self using functional MRI. Eleven healthy volunteers were scanned with echoplanar imaging using the blood oxygen level-dependent contrast method. The task consisted of aurally delivered statements requiring a yes-no decision. In the experimental condition, participants responded to a variety of statements requiring knowledge of and reflection on their own abilities, traits and attitudes (e.g. 'I forget important things', 'I'm a good friend', 'I have a quick temper'). In the control condition, participants responded to statements requiring a basic level of semantic knowledge (e.g. 'Ten seconds is more than a minute', 'You need water to live'). The latter condition was intended to control for auditory comprehension, attentional demands, decision-making, the motoric response, and any common retrieval processes. Individual analyses revealed consistent anterior medial prefrontal and posterior cingulate activation for all participants. The overall activity for the group, using a random-effects model, occurred in anterior medial prefrontal cortex (t = 13.0, corrected P = 0.05; x, y, z, 0, 54, 8, respectively) and the posterior cingulate (t = 14.7, P = 0.02; x, y, z, -2, -62, 32, respectively; 967 voxel extent). These data are consistent with lesion studies of impaired awareness, and suggest that the medial prefrontal and posterior cingulate cortex are part of a neural system subserving self-reflective thought.
---238---
Anesthesiology. 2000 May;92(5):1257-67.
Neural mechanisms of antinociceptive effects of hypnosis.
Faymonville ME, Laureys S, Degueldre C, DelFiore G, Luxen A, Franck G, Lamy M, Maquet P.
Departments of Anesthesiology and Intensive Care Medicine and Neurology, and the Cyclotron Research Centre, University Hospital of Liège, Liège, Belgium. anesrea@ulg.ac.be
BACKGROUND: The neural mechanisms underlying the modulation of pain perception by hypnosis remain obscure. In this study, we used positron emission tomography in 11 healthy volunteers to identify the brain areas in which hypnosis modulates cerebral responses to a noxious stimulus. METHODS: The protocol used a factorial design with two factors: state (hypnotic state, resting state, mental imagery) and stimulation (warm non-noxious vs. hot noxious stimuli applied to right thenar eminence). Two cerebral blood flow scans were obtained with the 15O-water technique during each condition. After each scan, the subject was asked to rate pain sensation and unpleasantness. Statistical parametric mapping was used to determine the main effects of noxious stimulation and hypnotic state as well as state-by-stimulation interactions (i.e., brain areas that would be more or less activated in hypnosis than in control conditions, under noxious stimulation). RESULTS: Hypnosis decreased both pain sensation and the unpleasantness of noxious stimuli. Noxious stimulation caused an increase in regional cerebral blood flow in the thalamic nuclei and anterior cingulate and insular cortices. The hypnotic state induced a significant activation of a right-sided extrastriate area and the anterior cingulate cortex. The interaction analysis showed that the activity in the anterior (mid-)cingulate cortex was related to pain perception and unpleasantness differently in the hypnotic state than in control situations. CONCLUSIONS: Both intensity and unpleasantness of the noxious stimuli are reduced during the hypnotic state. In addition, hypnotic modulation of pain is mediated by the anterior cingulate cortex.
---32---
Neuroreport. 1997 Jan 20;8(2):561-5.
FMRI of the prefrontal cortex during overt verbal fluency.
Phelps EA, Hyder F, Blamire AM, Shulman RG.
Department of Psychology, Yale University, New Haven, CT 06520, USA.
Verbal fluency is known to be associated with activity in the left prefrontal cortex. Recent positron emission tomography (PET) results confirmed this finding. In the present study, high resolution functional magnetic resonance imaging (fMRI) was used to further localize activity in the prefrontal cortex related to verbal fluency. Activation was observed in three behavioral tasks: (1) Repeat-subjects repeated words, (2) Opposite-subjects produced the antonym of words, and (3) Generate-subjects generated words beginning with a given letter. When comparing Generate with both Repeat and Opposite, we observed small areas of activation in the left inferior frontal gyrus and anterior cingulate, similar to the centers of mass reported using PET. We also found additional activation around the superior frontal sulcus.
-----278---
Science. 2003 Mar 21;299(5614):1864.
Imperceptible stimuli and sensory processing impediment.
Blankenburg F, Taskin B, Ruben J, Moosmann M, Ritter P, Curio G, Villringer A.
Decrease during imperceptible electric finger stimulation. Left index finger 7Hz electric pulse subliminal stimulation versus no stimulation. WOEXP: 278.
---86---
Neuroimage. 2001 Jan;13(1):143-52.
Cortical activations during the mental rotation of different visual objects.
Jordan K, Heinze HJ, Lutz K, Kanowski M, Jäncke L.
Institute of General Psychology, Otto-von-Guericke University Magdeburg, Magdeburg, D-39106, Germany.
Whole-head functional magnetic resonance imaging was applied to nine healthy right-handed subjects while they were performing three different mental rotation tasks and two visual control tasks. The mental rotation tasks comprised stimuli pairs derived from the "classical" 3D cube figures first used by R. N. Shepard and J. Metzler (1971, Science 171, 701-703), pairs of letters, and pairs of abstract figures developed by J. Hochberg and L. Gellmann (1977, Memory Cognit. 5, 23-26). In some cases, the paired objects were identical except that they were rotated in a certain plane. In other cases, the two objects were incongruent. Subjects were shown one pair of objects at a time and asked to judge whether the two were the same. In line with previous studies we found that decision times increased linearly with the degree of separation between the two objects. Cortical activation converged to demonstrate bilateral core regions in the superior and inferior parietal lobe (centered on the intraparietal sulcus), which were similarly activated during all three mental rotation tasks. Thus, our results suggest that different kinds of stimuli used for mental rotation tasks did not inevitably evoke activations outside the parietal core regions. For example we did not find any activation in brain areas known to be involved in lexical or verbal processing nor activations in cortical regions known to be involved in object identification or classification.
SPECIFICALLY Mental rotation of figures versus object determination or dots counting. Deciding whether visual stimuli were the same or mirrored indicating by pressing one of two buttons with the index or the middle finger of their right hand. WOEXP: 86.
---13---
Proc Natl Acad Sci U S A. 1997 Jun 24;94(13):6989-94.
"Willed action": a functional MRI study of the human prefrontal cortex during a sensorimotor task.
Hyder F, Phelps EA, Wiggins CJ, Labar KS, Blamire AM, Shulman RG.
Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT 06510, USA. hyder@mrcbs.med.yale.edu
Functional MRI (fMRI) was used to examine human brain activity within the dorsolateral prefrontal cortex during a sensorimotor task that had been proposed to require selection between several responses, a cognitive concept termed "willed action" in a positron emission tomography (PET) study by Frith et al. [Frith, C. D., Friston, K., Liddle, P. F. & Frackowiak, R. S. J. (1991) Proc. R. Soc. London Ser. B 244, 241-246]. We repeated their sensorimotor task, in which the subject chooses to move either of two fingers after a stimulus, by fMRI experiments in a 2.1-T imaging spectrometer. Echo-planar images were acquired from four coronal slices in the prefrontal cortex from nine healthy subjects. Slices were 5 mm thick, centers separated by 7 mm, with nominal in-plane spatial resolution of 9.6 x 5.0 mm2 for mean data. Our mean results are in agreement with the PET results in that we saw similar bilateral activations. The present results are compared with our previously published fMRI study of a verbal fluency task, which had also been proposed by Frith et al. to elicit a "willed action" response. We find a clear separation of activation foci in the left dorsolateral prefrontal cortex for the sensorimotor (Brodmann area 46) and verbal fluency (Brodmann area 45) tasks. Hence, assigning a particular activated region to "willed action" is not supported by the fMRI data when examined closely because identical regions are not activated with different modalities. Similar modality linked activations can be observed in the original PET study but the greater resolution of the fMRI data makes the modality linkages more definite.
--EGNER---
Egner, T., Hirsch, J. (2005). Cognitive control mechanisms resolve conflict
through cortical amplification of task-relevant information.
Nature Neuroscience, 8 (12), 1784-1790.
--101---
J Cogn Neurosci. 2000 Sep;12(5):763-74.
Brain activation during mental transformation of size.
Larsen A, Bundesen C, Kyllingsbaek S, Paulson OB, Law I.
Center for Visual Cognition, Department of Psychology, University of Copenhagen, Denmark.
Visual comparison between different-sized objects with respect to shape can be done by encoding one of the objects as a mental image, transforming the image to the size format of the other object, and then testing for a match (Bundesen, C., & Larsen, A. [1975]. Visual transformation of size. Journal of Experimental Psychology: Human Perception and Performance, 1, 214-220). To identify the brain structures implicated in mental transformation of size, we measured the distribution of regional cerebral blood flow (rCBF) by positron emission tomography (PET) in 12 normal subjects who compared random stimulus patterns with respect to shape regardless of variations in size in a one-back match-to-sample paradigm. Each subject was PET-scanned 12 times during repetitive injections of H(2)(15)O. In one condition (three scans), all stimulus patterns were small. In a second condition (three scans), all stimuli were large. In the third condition (six scans), the stimuli alternated between small and large. Mental transformation of size should occur in the alternating-size condition but not in the fixed-size conditions. As expected, behavioral measures (reaction time [RT], d', beta) were nearly the same for the two fixed-size conditions but mean RT was longer and d' smaller in the alternating-size condition. Changes in rCBF specific to mental transformation of size were estimated by contrasting the alternating-size with the fixed-size conditions by use of statistical parametric mapping (SPM96) at a threshold of p <. 05 corrected for multiple comparisons. The detected brain structures implicated in mental transformation of size were primarily located in the dorsal pathways, comprising structures in the occipital, parietal, and temporal transition zone (predominantly in the left hemisphere), posterior parietal cortex (bilaterally), area MT/V5 (left), and vermis (bilaterally). Contrasts between the two fixed-size conditions showed significant effects in only the occipital cortex.
Large line patterns. One-back match-to-sample task with large line patterns versus small line patterns. WOEXP: 101.
--23---previous
---261---
Negative interaction between predictable tones and button press. Negative interaction between predictable tones and self-paced button presses versus no button presses and random tones with button press. WOEXP: 261.
Neuropsychologia. 1998 Jun;36(6):521-9.
How do we predict the consequences of our actions? A functional imaging study.
Blakemore SJ, Rees G, Frith CD.
Wellcome Department of Cognitive Neurology, Institute of Neurology, London. s.blakemore@ucl.ac.uk
Humans are readily able to distinguish expected and unexpected sensory events. Whether a single mechanism underlies this ability is unknown. The most common type of expected sensory events are those generated as a consequence of self-generated actions. Using H2 15O PET, we studied brain responses to such predictable sensory events (tones) and to similar unpredictable events and especially how the processing of predictable sensory events is modified by the context of a causative self-generated action. Increases in activity when the tones were unpredictable were seen in the inferior and superior temporal lobe bilaterally, the right parahippocampal gyrus and right parietal cortex. Self-generated actions produced activity in a number of motor and premotor areas, including dorsolateral prefrontal cortex. We observed an interaction between the predictability of stimuli and self-generated actions in several areas, including the medial posterior cingulate cortex, left insula, dorsomedial thalamus, superior colliculus and right inferior temporal cortex. This modulation of activity associated with stimulus predictability in the context of self-generated actions implies that these areas may be involved in self-monitoring processes. Detection of expected stimuli and the detection of the sensory consequences of self-generated actions appear to be functionally distinct processes, and are carried out in different cortical areas. These observations support theoretical approaches to cognition that postulate the existence of a self-monitoring system.
---185---
Nature. 2001 Jun 21;411(6840):950-3.
Spatial awareness is a function of the temporal not the posterior parietal lobe.
Karnath HO, Ferber S, Himmelbach M.
Department of Cognitive Neurology, University of Tübingen, Germany. karnath@uni-tuebingen.de
Comment in:
Nature. 2001 Jun 21;411(6840):903-4.
Our current understanding of spatial behaviour and parietal lobe function is largely based on the belief that spatial neglect in humans (a lack of awareness of space on the side of the body contralateral to a brain injury) is typically associated with lesions of the posterior parietal lobe. However, in monkeys, this disorder is observed after lesions of the superior temporal cortex, a puzzling discrepancy between the species. Here we show that, contrary to the widely accepted view, the superior temporal cortex is the neural substrate of spatial neglect in humans, as it is in monkeys. Unlike the monkey brain, spatial awareness in humans is a function largely confined to the right superior temporal cortex, a location topographically reminiscent of that for language on the left. Hence, the decisive phylogenetic transition from monkey to human brain seems to be a restriction of a formerly bilateral function to the right side, rather than a shift from the temporal to the parietal lobe. One may speculate that this lateralization of spatial awareness parallels the emergence of an elaborate representation for language on the left side.
Patients with spatial neglect and right brain damage from infarct or hemorrhage versus right brain damage patients without spatial neglect. WOEXP: 185.
---565---
Proc Natl Acad Sci U S A. 1999 Feb 16;96(4):1794-9.
Task-related and item-related brain processes of memory retrieval.
Düzel E, Cabeza R, Picton TW, Yonelinas AP, Scheich H, Heinze HJ, Tulving E.
Department of Neurology II, Otto von Guericke University of Magdeburg, Leipziger Strasse 44, 39120 Magdeburg, Germany. emrah.duezel@medizin.uni-magdeburg.de
In all cognitive tasks, general task-related processes operate throughout a given task on all items, whereas specific item-related processes operate differentially on individual items. In typical functional neuroimaging experiments, these two sets of processes have usually been confounded. Herein we report a combined positron emission tomography and event-related potential (ERP) experiment that was designed to distinguish between neural correlates of task-related and item-related processes of memory retrieval. Two retrieval tasks, episodic and semantic, were crossed with episodic (old/new) and semantic (living/nonliving) properties of individual items to yield evidence of regional brain activity associated with task-related processes, item-related processes, and their interaction. The results showed that episodic retrieval task was associated with increased blood flow in right prefrontal and posterior cingulate cortex, as well as with a sustained right-frontopolar-positive ERP, but that the semantic retrieval task was associated with left frontal and temporal lobe activity. Retrieval of old items was associated with increased blood flow in the left medial temporal lobe and with a brief late positive ERP component. The results provide converging hemodynamic and electrophysiological evidence for the distinction of task- and item-related processes, show that they map onto spatially and temporally distinct patterns of brain activity, and clarify the hemispheric encoding/retrieval asymmetry (HERA) model of prefrontal encoding and retrieval asymmetry.
Task-related episodic retrieval versus semantic. Episodic retrieval with a decision whether a visually presented word was presented in an encoding list with right hand button response versus semantic retrieval. WOEXP: 565.
---248---
Ann Neurol. 1999 Jan;45(1):40-7.
Region-specific encoding of sensory and affective components of pain in the human brain: a positron emission tomography correlation analysis.
Tölle TR, Kaufmann T, Siessmeier T, Lautenbacher S, Berthele A, Munz F, Zieglgänsberger W, Willoch F, Schwaiger M, Conrad B, Bartenstein P.
Department of Neurology, Technical University, Munich, Germany.
Brain imaging with positron emission tomography has identified some of the principal cerebral structures of a central network activated by pain. To discover whether the different cortical and subcortical areas process different components of the multidimensional nature of pain, we performed a regression analysis between noxious heat-related regional blood flow increases and experimental pain parameters reflecting detection of pain, encoding of pain intensity, as well as pain unpleasantness. The results of our activation study indicate that different functions in pain processing can be attributed to different brain regions; ie, the gating function reflected by the pain threshold appeared to be related to anterior cingulate cortex, the frontal inferior cortex, and the thalamus, the coding of pain intensity to the periventricular gray as well as to the posterior cingulate cortex, and the encoding of pain unpleasantness to the posterior sector of the anterior cingulate cortex.
Correlation with pain intensity. Correlation with subjective ratings of pain intensity with hot pain right volar forearm. WOEXP: 248.
--363---
J Neurosci. 1994 Jul;14(7):4095-108.
Distributed processing of pain and vibration by the human brain.
Coghill RC, Talbot JD, Evans AC, Meyer E, Gjedde A, Bushnell MC, Duncan GH.
Centre de Recherche en Sciences Neurologiques, Université de Montréal, Quebec, Canada.
Pain is a diverse sensory and emotional experience that likely involves activation of numerous regions of the brain. Yet, many of these areas are also implicated in the processing of nonpainful somatosensory information. In order to better characterize the processing of pain within the human brain, activation produced by noxious stimuli was compared with that produced by robust innocuous stimuli. Painful heat (47-48 degrees C), nonpainful vibratory (110 Hz), and neutral control (34 degrees C) stimuli were applied to the left forearm of right-handed male subjects. Activation of regions within the diencephalon and telencephalon was evaluated by measuring regional cerebral blood flow using positron emission tomography (15O-water-bolus method). Painful stimulation produced contralateral activation in primary and secondary somatosensory cortices (SI and SII), anterior cingulate cortex, anterior insula, the supplemental motor area of the frontal cortex, and thalamus. Vibrotactile stimulation produced activation in contralateral SI, and bilaterally in SII and posterior insular cortices. A direct comparison of pain and vibrotactile stimulation revealed that both stimuli produced activation in similar regions of SI and SII, regions long thought to be involved in basic somatosensory processing. In contrast, painful stimuli were significantly more effective in activating the anterior insula, a region heavily linked with both somatosensory and limbic systems. Such connections may provide one route through which nociceptive input may be integrated with memory in order to allow a full appreciation of the meaning and dangers of painful stimuli. These data reveal that pain-related activation, although predominantly contralateral in distribution, is more widely dispersed across both cortical and thalamic regions than that produced during innocuous vibrotactile stimulation. This distributed cerebral activation reflects the complex nature of pain, involving discriminative, affective, autonomic, and motoric components. Furthermore, the high degree of interconnectivity among activated regions may account for the difficulty of eliminating pathological pain with discrete CNS lesions.
---39---
J Cogn Neurosci. 2000 Sep;12(5):711-20.
Brain areas involved in perception of biological motion.
Grossman E, Donnelly M, Price R, Pickens D, Morgan V, Neighbor G, Blake R.
Department of Psychology, Vanderbilt University, Nashville, TN 37240, USA. e.grossman@vanderbilt.edu
These experiments use functional magnetic resonance imaging (fMRI) to reveal neural activity uniquely associated with perception of biological motion. We isolated brain areas activated during the viewing of point-light figures, then compared those areas to regions known to be involved in coherent-motion perception and kinetic-boundary perception. Coherent motion activated a region matching previous reports of human MT/MST complex located on the temporo-parieto-occipital junction. Kinetic boundaries activated a region posterior and adjacent to human MT previously identified as the kinetic-occipital (KO) region or the lateral-occipital (LO) complex. The pattern of activation during viewing of biological motion was located within a small region on the ventral bank of the occipital extent of the superior-temporal sulcus (STS). This region is located lateral and anterior to human MT/MST, and anterior to KO. Among our observers, we localized this region more frequently in the right hemisphere than in the left. This was true regardless of whether the point-light figures were presented in the right or left hemifield. A small region in the medial cerebellum was also active when observers viewed biological-motion sequences. Consistent with earlier neuroimaging and single-unit studies, this pattern of results points to the existence of neural mechanisms specialized for analysis of the kinematics defining biological motion.
Biological visual motion. Biological motion of dots versus scrambled motion of dots. WOEXP: 111.
----41---
Nat Neurosci. 2000 Mar;3(3):277-83.
A multimodal cortical network for the detection of changes in the sensory environment.
Downar J, Crawley AP, Mikulis DJ, Davis KD.
Institute of Medical Science, University of Toronto, and Toronto Western Research Institute, MP14-322, 399 Bathurst Street, Toronto, Ontario, M5T 2S8, Canada.
Sensory stimuli undergoing sudden changes draw attention and preferentially enter our awareness. We used event-related functional magnetic-resonance imaging (fMRI) to identify brain regions responsive to changes in visual, auditory and tactile stimuli. Unimodally responsive areas included visual, auditory and somatosensory association cortex. Multimodally responsive areas comprised a right-lateralized network including the temporoparietal junction, inferior frontal gyrus, insula and left cingulate and supplementary motor areas. These results reveal a distributed, multimodal network for involuntary attention to events in the sensory environment. This network contains areas thought to underlie the P300 event-related potential and closely corresponds to the set of cortical regions damaged in patients with hemineglect syndromes.
ENVIRONMENTAL SOUNDS Auditory change. Change between two sounds, running water and croaking frogs versus change in visual or tactile stimuli. WOEXP: 454.
---22---
Hum Brain Mapp. 2000 Jun;10(2):74-9.
Lateralized automatic auditory processing of phonetic versus musical information: a PET study.
Tervaniemi M, Medvedev SV, Alho K, Pakhomov SV, Roudas MS, Van Zuijen TL, Näätänen R.
Cognitive Brain Research Unit, Department of Psychology, University of Helsinki, Finland. Tervanie@Helsinki.Fi
Previous positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) studies show that during attentive listening, processing of phonetic information is associated with higher activity in the left auditory cortex than in the right auditory cortex while the opposite is true for musical information. The present PET study determined whether automatically activated neural mechanisms for phonetic and musical information are lateralized. To this end, subjects engaged in a visual word classification task were presented with phonetic sound sequences consisting of frequent (P = 0.8) and infrequent (P = 0.2) phonemes and with musical sound sequences consisting of frequent (P = 0.8) and infrequent (P = 0.2) chords. The phonemes and chords were matched in spectral complexity as well as in the magnitude of frequency difference between the frequent and infrequent sounds (/e/ vs. /o/; A major vs. A minor). In addition, control sequences, consisting of either frequent (/e/; A major) or infrequent sounds (/o/; A minor) were employed in separate blocks. When sound sequences consisted of intermixed frequent and infrequent sounds, automatic phonetic processing was lateralized to the left hemisphere and musical to the right hemisphere. This lateralization, however, did not occur in control blocks with one type of sound (frequent or infrequent). The data thus indicate that automatic activation of lateralized neuronal circuits requires sound comparison based on short-term sound representations.
Chords simulation: standard sequence versus deviant sequence. WOEXP: 22.
---134--
J Cogn Neurosci. 2000;12 Suppl 2:118-29.
Neural activation during response competition.
Hazeltine E, Poldrack R, Gabrieli JD.
NASA Ames Research Center, Moffett Field, CA 94305, USA.
The flanker task, introduced by Eriksen and Eriksen [Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16, 143--149], provides a means to selectively manipulate the presence or absence of response competition while keeping other task demands constant. We measured brain activity using functional magnetic resonance imaging (fMRI) during performance of the flanker task. In accordance with previous behavioral studies, trials in which the flanking stimuli indicated a different response than the central stimulus were performed significantly more slowly than trials in which all the stimuli indicated the same response. This reaction time effect was accompanied by increases in activity in four regions: the right ventrolateral prefrontal cortex, the supplementary motor area, the left superior parietal lobe, and the left anterior parietal cortex. The increases were not due to changes in stimulus complexity or the need to overcome previously learned associations between stimuli and responses. Correspondences between this study and other experiments manipulating response interference suggest that the frontal foci may be related to response inhibition processes whereas the posterior foci may be related to the activation of representations of the inappropriate responses.
Response competition. Visual presentation of three colored circles with response by pressing of either of two buttons determined by the color of the center circle. Incongruent trials with flanking circles indicating a competing response versus congruent trials with flanking circles indicating the same response as the center circle. WOEXP: 134.
----365---
J Neurosci. 1994 Jul;14(7):4095-108.
Distributed processing of pain and vibration by the human brain.
Coghill RC, Talbot JD, Evans AC, Meyer E, Gjedde A, Bushnell MC, Duncan GH.
Centre de Recherche en Sciences Neurologiques, Université de Montréal, Quebec, Canada.
Pain is a diverse sensory and emotional experience that likely involves activation of numerous regions of the brain. Yet, many of these areas are also implicated in the processing of nonpainful somatosensory information. In order to better characterize the processing of pain within the human brain, activation produced by noxious stimuli was compared with that produced by robust innocuous stimuli. Painful heat (47-48 degrees C), nonpainful vibratory (110 Hz), and neutral control (34 degrees C) stimuli were applied to the left forearm of right-handed male subjects. Activation of regions within the diencephalon and telencephalon was evaluated by measuring regional cerebral blood flow using positron emission tomography (15O-water-bolus method). Painful stimulation produced contralateral activation in primary and secondary somatosensory cortices (SI and SII), anterior cingulate cortex, anterior insula, the supplemental motor area of the frontal cortex, and thalamus. Vibrotactile stimulation produced activation in contralateral SI, and bilaterally in SII and posterior insular cortices. A direct comparison of pain and vibrotactile stimulation revealed that both stimuli produced activation in similar regions of SI and SII, regions long thought to be involved in basic somatosensory processing. In contrast, painful stimuli were significantly more effective in activating the anterior insula, a region heavily linked with both somatosensory and limbic systems. Such connections may provide one route through which nociceptive input may be integrated with memory in order to allow a full appreciation of the meaning and dangers of painful stimuli. These data reveal that pain-related activation, although predominantly contralateral in distribution, is more widely dispersed across both cortical and thalamic regions than that produced during innocuous vibrotactile stimulation. This distributed cerebral activation reflects the complex nature of pain, involving discriminative, affective, autonomic, and motoric components. Furthermore, the high degree of interconnectivity among activated regions may account for the difficulty of eliminating pathological pain with discrete CNS lesions.
----534---
Neuroreport. 2005 Apr 25;16(6):649-52.
Motion verb sentences activate left posterior middle temporal cortex despite static context.
Wallentin M, Lund TE, Ostergaard S, Ostergaard L, Roepstorff A.
Center for Semiotics, Aarhus University, Niels Juels Gade 84, 8200 Arhus N, Denmark. mikkel@pet.auh.dk
The left posterior middle temporal region, anterior to V5/MT, has been shown to be responsive both to images with implied motion, to simulated motion, and to motion verbs. In this study, we investigated whether sentence context alters the response of the left posterior middle temporal region. 'Fictive motion' sentences are sentences in which an inanimate subject noun, semantically incapable of self movement, is coupled with a motion verb, yielding an apparent semantic contradiction (e.g. 'The path comes into the garden.'). However, this context yields no less activation in the left posterior middle temporal region than sentences in which the motion can be applied to the subject noun. We speculate that the left posterior middle temporal region activity in fictive motion sentences reflects the fact that the hearer applies motion to the depicted scenario by scanning it egocentrically.
The left posterior middle temporal region, anterior to V5/MT, has been shown to be responsive both to images with implied motion, to simulated motion, and to motion verbs

 浙公网安备 33010602011771号
浙公网安备 33010602011771号