Chapter15 Principles of Metabolic Regulation
(1)细胞和有机体维持着动态稳定
- 调节作用的基因多 2500个 12%
- 激酶占比多 300个
- 一个通路受到多方调节
(2)酶的数量和催化活性 可以被调节
- 反应物的调控
- 糖酵解的产物,代谢物和cofactor,底物
- 信号分子的调控
- cAMP,Ca2+
①细胞外信号:激素、神经信号(乙酰胆碱),生长因子,细胞因子
- 调节转录因子的结合
- 相关功能的酶共享response element,被一同调节
②mRNA
③蛋白合成后的调控
- 蛋白可以被泛素识别降解
- 快速关闭某一蛋白的作用很费能量,但是有助于机体快速响应
④转运蛋白
- 细胞质被分割成小区室依靠膜上转运到白调控反应物浓度
代谢组学的研究工具:DNA微阵列、二维凝胶电泳
一旦酶被合成,如何调控:
- 底物的浓度
- 变构影响因子
- 共价修饰
- 磷酸化或去磷酸化
- 阻遏因子的去除
- 结合其他因子
- PKA的激活要cAMP的结合
(3)细胞内有些反应是近乎平衡的,在底物浓度的变化下反应反向会变化
还有一些反应处在非平衡状态,他们是调控的位点
- 以ATP反应供能为例,如果反应平衡了,ATP就不再有正反应的驱动力
- 所以机体要有敏感的调控确保ATP浓度远高于平衡浓度
- 机体的代谢通路中的关键反应都是极度不平衡的,且受到多方调控(PFK-1的反应)
(4)腺嘌呤核苷酸在代谢调控中起到重要作用(AMPK维持ATP足量)
- ATP/AMP 含量是细胞能量状况的标志
- ATP的缺少使得ATP参与反应的酶 无法结合足够的底物(ATP)
- 热力学上ATP维持高浓度推动需要ATP的反应正向进行
- 当饥饿时,ATP会减少,AMP会增多:
- 机体AMP变构 AMP蛋白激酶AMPK ,使其被LKB1磷酸化激活
- AMPK激活后推动糖酵解,脂肪酸氧化,葡萄糖转运;抑制糖原合成
(5)除了ATP,其他物质在体内也要维持稳定:NADH、NADPH、葡萄糖
- 脑细胞没有储糖能力,全靠胰岛素和胰高血糖素(glucagon)来平衡脑补供糖
- 进化的压力使得调节机制处于如下协调的状态:
①最大化能量利用效率(不同时做反方向的事)
②合理分配主干和支路代谢途径
③选择合适的供能物质
④在产物聚集时,减少合成
15.2 代谢控制分析
(1)三个参数
① 通量控制系数 C
- 代谢途径中每个酶对调节代谢物流速的相对贡献
② 弹性系数 ε
- 单个酶对代谢物或效应物浓度的反应性,是酶固有的动力学性质
![]()

③反应系数 R
- 外部控制物对流经代谢途径通量的影响
总和原理 代谢途径各步反应C之和为1
参数相关原理 R = C·ε
(2)control 和 regulation的概念
- control 作用的酶是调节靶点,调控代谢的通路
- 增加可以使代谢加倍
- regulation 作用的酶是维持机体的稳态,不至于内环境改变造成代谢突然变化
- 增加不能使代谢加倍
(3)可以通过提高某一通路的所有酶浓度来提高一条路的flux
15.3 糖酵解和糖异生的协同调控
- substrate cycle 底物循环虽然额外地消耗了能量,但是对机体调控有利
(1)己糖激酶的同工酶在肌肉和肝脏中以不同方式调节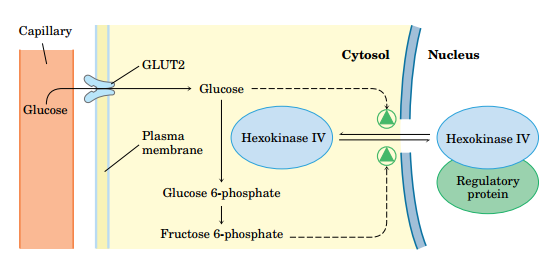
- 肝中的己糖激酶IV(葡糖激酶)
- Km值大,
- 不被催化产物G6P抑制
- 调控方式:
- 高血糖时,葡萄糖通过GLUT2转运入肝细胞,被高效合成糖原
- 当血糖浓度降低时,F6P刺激调控蛋白与酶IV的结合(与细胞核内),抑制反应
- 葡萄糖和F6P竞争结合,当葡萄糖含量升高,酶IV被释放
- 一般的己糖激酶在产物G6P升高时会被抑制
(2)己糖激酶IV 和 G6Pase 可以在转录水平调节
(3)PFK-1 和 F1,6BPase 是互为相反调节的
- 糖酵解、柠檬酸产物堆积
- AMP、ADP堆积
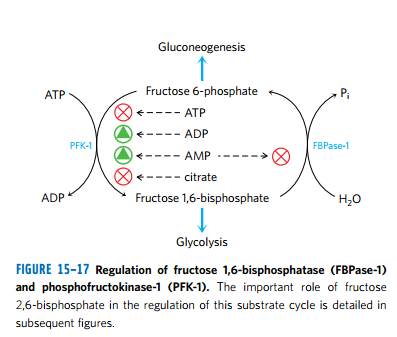
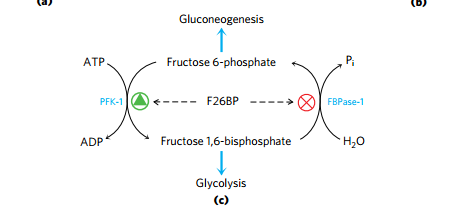
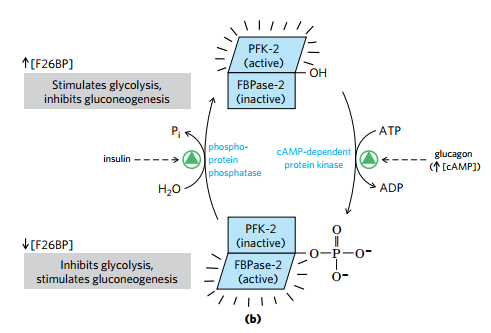
(4)F2,6BP 是强有力的PFK-1、FBPase-1 别构调节剂
- PFK-2/FPBase-2 是同一个蛋白不同结构域
- 胰岛素和胰高血糖素通过对 PFK-2/FPBase-2 磷酸化情况的控制,控制F2,6BP的含量控制PFK-1、FBPase-1以决定糖酵解糖异生方向
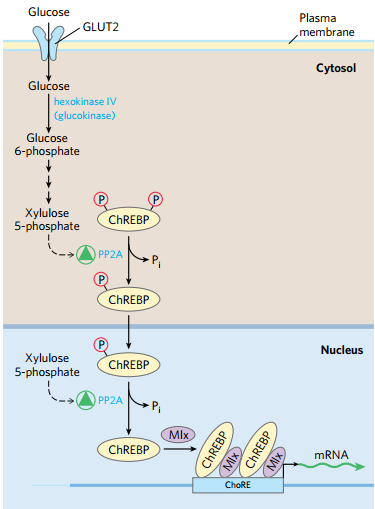
(5)木酮糖对于糖脂肪代谢是一个关键调控物
- 木酮糖由戊糖途径生成
- 磷酸化激活PP2A
- PP2A 脱氢激活PFK-2,F2,6BP升高
- 增加了乙酰CoA的生成和NADPH的生成
(5)丙酮酸激酶 的调控
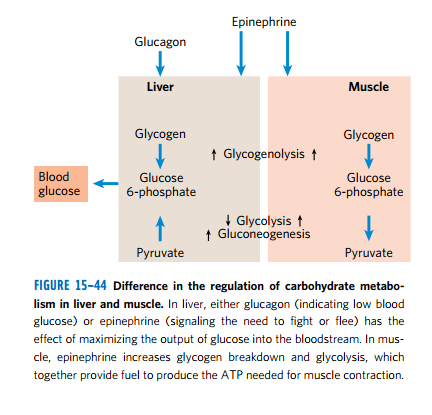
- 在肝脏中,血糖降低使得胰高血糖素分泌,通过cAMP通路失活丙酮酸激酶,使得更过糖流向别处
- 而在肌肉,肾上腺素刺激的cAMP上升会刺激糖原的分解和糖酵解来满足所需
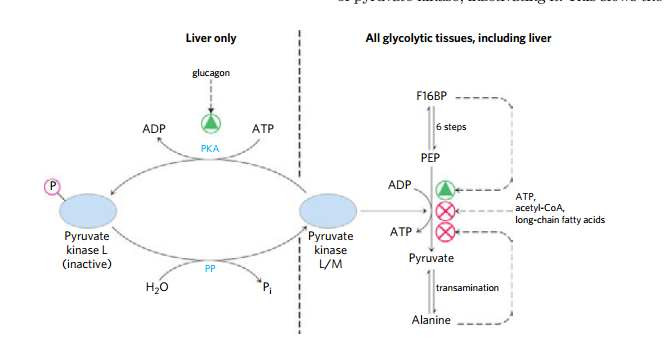
(6) 糖异生丙酮酸羧激酶的调节

(7)对糖酵解、糖异生 的转录水平调节改变酶的数量
① 转录因子:ChREBP糖相应元件结合蛋白
- 戊糖途径生成的木酮糖xylulose给机体带来了糖酵解底物充足的信号,诱生PP2A使得ChRBP去磷酸化
- ChRBP作为转录因子结合基因上游
② 转录因子:SREBP-1c 固醇调控元件结合蛋白
- 受胰岛素刺激,促进脂肪合成相关转录
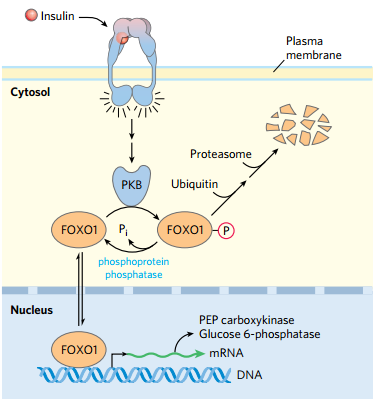
③转录因子:CREB 环腺苷磷酸响应原件结合蛋白
- FOXO1 是一种CREB
- 胰高血糖素可以阻止PKB对FOXO1的磷酸化,使其促进转录糖异生相关蛋白
15.3 SUMMARY
- 糖酵解和糖异生有3个不共享酶的反应,这是调控的位点
- 己糖激酶在肝中有特殊作用:低血糖时释放葡萄糖入血,高血糖时合成糖原
- PFK-1被ATP别构抑制,F2,6BP别构激活
- 丙酮酸激酶被ATP别构抑制,在肝中还能被cAMP磷酸化而抑制
- 糖再生的调控位点在丙酮酸羧化酶、FBPase-1
- F2,6BP对PFK-1,FBPase-1x的相反作用控制了糖酵解和糖异生的底物循环
- 胰高血糖素和肾上腺素通过cAMP磷酸化PFK-2/FBPase-2来减少F2,6BP;胰岛素的作用是去磷酸化
- 木酮糖作为一种信号促进糖酵解
- 转录因子接受外来信号,作用于DNA来调控酶合成
15.4 糖原的代谢
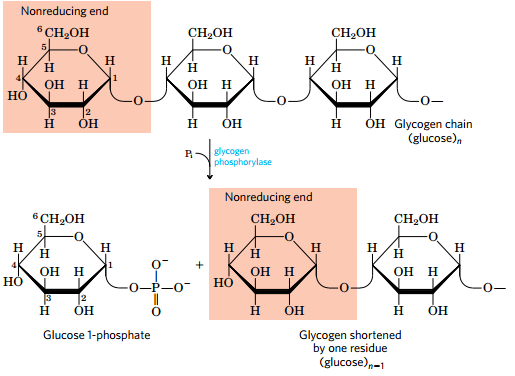
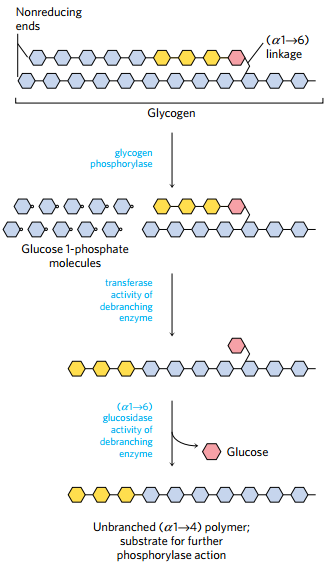
(1)糖原分解需要的酶
- 糖原磷酸化酶
- glycogen debranching enzyme
- 磷酸化 glucomutase
- 糖原分解产生的G1P由glucomutase变成G6P进入血液循环
(2)糖原的合成要让糖结合形成G-UDP,才能加到糖链上
- 这个结合是不可逆的,推动反应正向进行
- 尽管三磷酸核苷与葡糖磷酸反应的自由能小,但生成的焦磷酸水解推动反应
- 核苷对酶的非共价促进作用
- 对要进行糖原生成的糖分子进行了标记
- 糖原合成到一定长度要支化,形成的非还原糖基方便合成和分解糖原
![]()

(3)糖原合成的起始阶段需要引物
- 由glycogenin催化在自身合成的8bp糖链形成

15.5 糖原分解与合成的协同调控
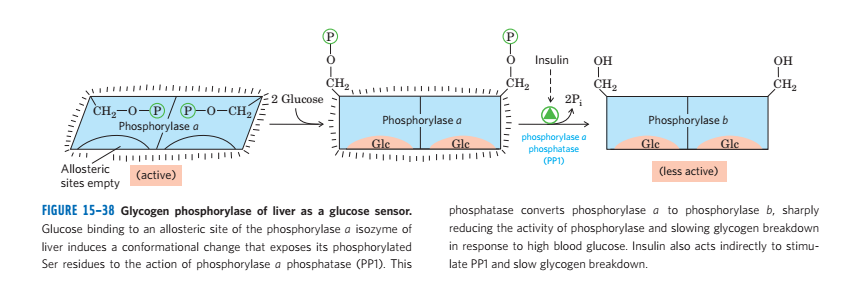
(1)糖原磷酸化酶受到变构调节和激素调节
- 通过一系列酶的影响,最终通过磷酸化的状态决定活性(磷酸化有活性)
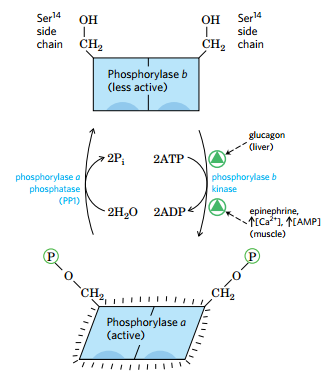
- 肾上腺素造成的级联反应
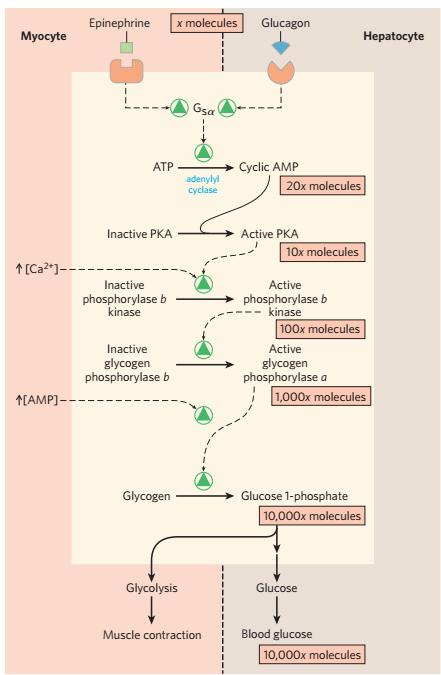
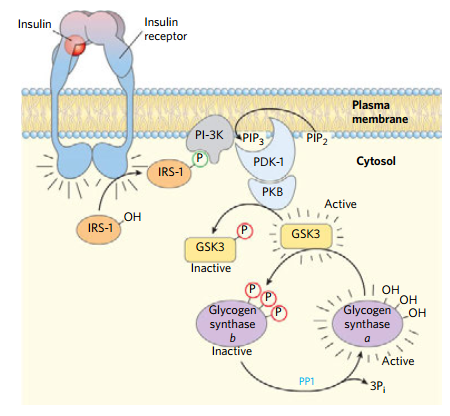
- 胰岛素通过去磷酸化降低酶活性
- 葡萄糖的增多别构抑制酶的活性
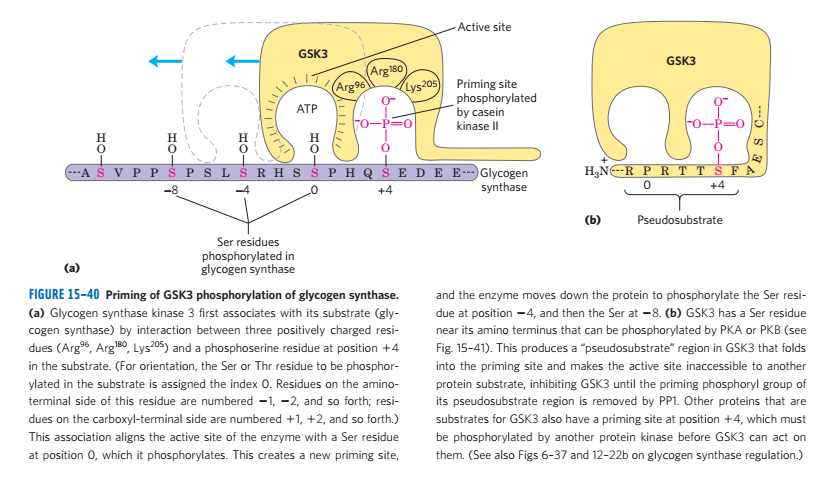
(2)糖原合成酶去磷酸化时表现出活性
- G6P进行变构调节
- 糖原合成酶去磷酸化状态有活性
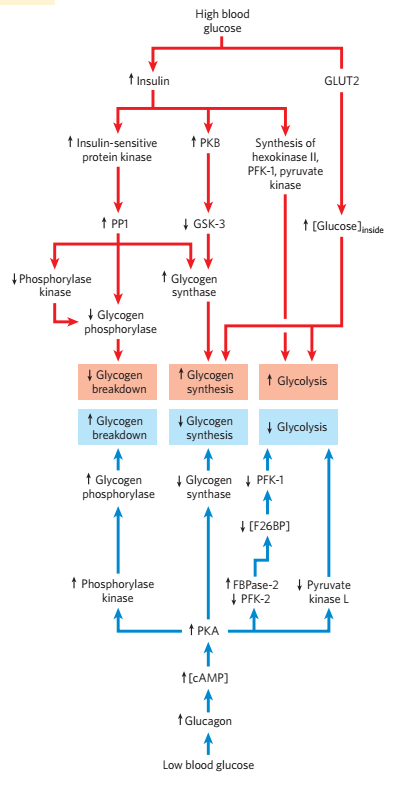
- 总之,胰高血糖素和肾上腺素造成糖原合成酶的磷酸化,和糖原磷酸化酶的磷酸化,增高血糖
- 胰岛素的作用相反

posted on 2018-11-13 22:10 iojafekniewg 阅读(648) 评论(0) 收藏 举报


 浙公网安备 33010602011771号
浙公网安备 33010602011771号