Chapter7 抑癌基因
一、实验证明,如果肿瘤不是肿瘤病毒产生时,与正常细胞融合后,其恶性表型是隐形的
二、
家族性视网膜母细胞瘤的形成模型
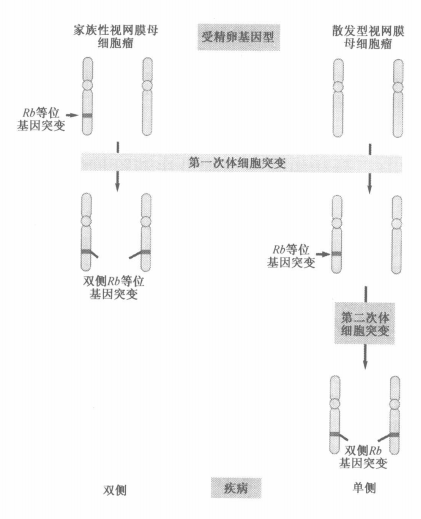
如何使得一个细胞获得两个突变(两次随机的突变可能性太小)
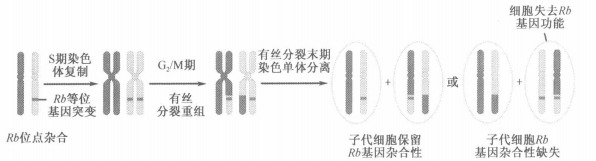
- 模型一:有丝分裂的同源重组
- 近亲结婚容易发生有丝分裂同源重组,导致杂合性缺失
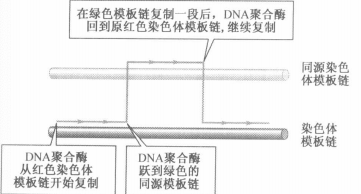
- 模型二:基因延长的时候聚合酶短暂地以同源染色体模板链进行合成
- 染色质部分丢失:有丝分裂时染色体不合理分离
三、通过杂合性缺失事件寻找抑癌基因
- RFLP 限制性片段长度多态性
- 聚合酶链式反应
寻找肿瘤细胞
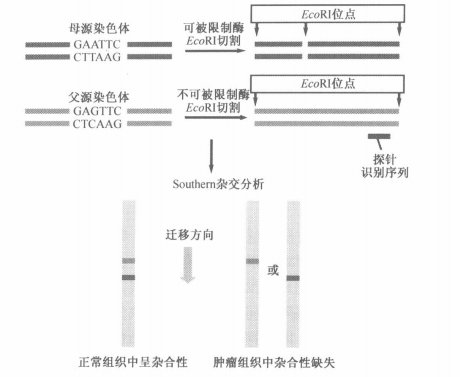
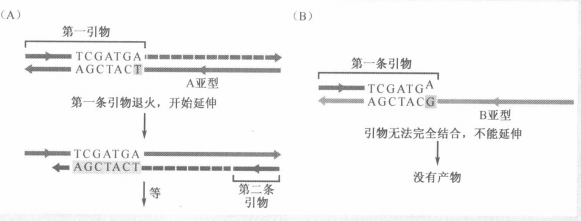
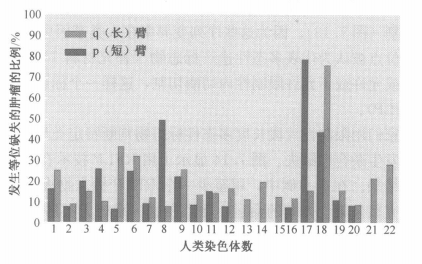
由此发现在癌细胞中,缺失率特别高的,很有可能含有抑癌基因
四、看门基因和看护基因
- 看门基因:
- 看护基因:维持基因组DNA序列的完整性
五、启动子甲基化是一种抑癌基因失活的重要形式
- 位于鸟嘌呤5'端的胞嘧啶甲基化可以下调这一区域基因的活性:MeCpG
- 导致组蛋白去乙酰化
- 这种甲基化还能被维持甲基化酶维持
- 在一些肿瘤发生进程中,会发生基因组甲基化水平进行性减少
![]()
- 导致高度重复序列的甲基化不足,染色体不稳定
- 高密度甲基化区域CpG岛在肿瘤基因组中常异常甲基化
- CpG岛常位于启动子区。导致一些基因沉默
- LOH(等位缺失)是家族性癌症的原因,而异常的甲基化是散发癌症的原因
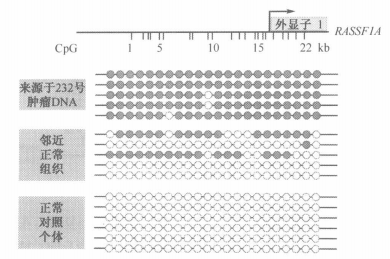
维甲酸 和 TSA 共用也许是乳腺癌治疗的好方法
维甲酸是一种细胞周期俘获剂,甚至可以促进细胞分化
但是乳腺癌细胞编码维甲酸受体的RARβ2基因被甲基化失活
组蛋白去乙酰化酶(HDAC)抑制剂TSA可以维持维甲酸受体的表达
六、抑癌基因通过多种方式起作用
- 直接抑制增殖:通过一系列生长抑制、分化诱导信号
- 控制细胞周期组分:代谢失衡、基因组损伤
- 还能编码肿瘤抑制基因
七、NF1蛋白作为一个Ras信号的负调控因子
- NF1与酵母的Ras GTPase激活蛋白IRA有很强的序列相关性
- 当有一个NF1等位基因突变时,NF1蛋白表达会减少,Ras活性会增高
- 单倍剂量不足
八、APC在结肠癌发病中的影响
肠壁上,干细胞位于肠隐窝内,受到周围细胞分泌的wnt刺激而增殖。然后在APC的作用下向外迁移,缺少了wnt信号,β-catemin也被降解,于是成为了成熟的肠壁细胞
- APC参与形成复合体,降解β-catenin
- 结肠癌细胞中APC蛋白翻译提前终止、或者启动子被甲基化而抑制转录
- 细胞获得干性,聚集在肠隐窝
- 经历了K-ras突变后,更具侵略性
- APC还与有丝分裂后期末期染色质分离有关,APC的缺失导致核型改变
九、pVHL调节缺氧翻译:脑视网膜血管瘤
- 正常氧压下pVHL蛋白使得HIF-1蛋白无效循环
- 低氧下HIF-1将不被降解
- HIF-1与 新血管生成、红细胞生成、糖代谢有关
- HIF-1的被pVHL识别依赖于HIP-1上的氧化羟脯氨酸,这与氧含量有关
- 有活性的HIF-1启动一系列基因:VEGF、PDGF、TGF-alpha的表达,导致很多细胞增殖
posted on 2018-11-28 22:06 iojafekniewg 阅读(895) 评论(0) 收藏 举报


 浙公网安备 33010602011771号
浙公网安备 33010602011771号