
非肿瘤生信:结肠靶向eNAMPT特异性肽系统用于治疗DSS诱导的小鼠急性和慢性结肠炎
题目
Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse
结肠靶向eNAMPT特异性肽系统用于治疗DSS诱导的小鼠急性和慢性结肠炎
Received: 2022.10.05 | Accepted: 2022.11.29 | Published: 2022.11.30
期刊名称:antioxidants
2021年影响因子 :7.675

摘要


Nicotinamide phosphoribosyl transferase (NAMPT) is required to maintain the NAD+ pool, among which extracellular (e) NAMPT is associated with inflammation, mainly mediated by macrophages. However, the role of (e) NAMPT in inflammatory macrophages in ulcerative colitis is insufficiently understood. Here our analyses of single-cell RNA-seq data revealed that the levels of NAMPT and CYBB/NOX2 in macrophages were elevated in patients with colitis and in mouse models of acute and chronic colitis. These findings indicate the clinical significance of NAMPT and CYBB in colitis. Further, we found that eNAMPT directly binds the extracellular domains of CYBB and TLR4 in activated NLRP3 inflammasomes. Moreover, we developed a recombinant 12-residue TK peptide designated colon-targeted (CT)-conjugated multifunctional NAMPT (rCTNAMPT), comprising CT as the colon-targeting moiety, which harbors the minimal essential residues required for CYBB/TLR4 binding. rCT-NAMPT effectively suppressed the severity of disease in DSSinduced acute and chronic colitis models through targeting the colon and inhibiting the interaction of NAMPT with CYBB or TLR4. Together, our data show that rCT-NAMPT may serve as an effective novel candidate therapeutic for colitis by modulating the NLRP3 inflammasome-mediated immune signaling system.
摘要:需要烟酰胺磷酸核糖基转移酶 (NAMPT) 来维持 NAD+ 池,其中细胞外 (e) NAMPT 与炎症相关,主要由巨噬细胞介导。然而,(e) NAMPT 在溃疡性结肠炎炎性巨噬细胞中的作用尚不清楚。在这里,我们对单细胞 RNA-seq 数据的分析表明,在结肠炎患者和急性和慢性结肠炎小鼠模型中,巨噬细胞中的 NAMPT 和 CYBB/NOX2 水平升高。这些发现表明 NAMPT 和 CYBB 在结肠炎中的临床意义。此外,我们发现 eNAMPT 直接结合激活的 NLRP3 炎性体中 CYBB 和 TLR4 的细胞外结构域。此外,我们开发了一种重组的 12 残基 TK 肽,命名为结肠靶向 (CT) 偶联的多功能 NAMPT (rCTNAMPT),其中包含 CT 作为结肠靶向部分,它含有 CYBB/TLR4 结合所需的最少必需残基。 rCT-NAMPT 通过靶向结肠并抑制 NAMPT 与 CYBB 或 TLR4 的相互作用,有效抑制了 DSS 诱导的急性和慢性结肠炎模型中疾病的严重程度。总之,我们的数据表明,rCT-NAMPT 可以通过调节 NLRP3 炎性体介导的免疫信号系统,作为结肠炎的一种有效的新型候选治疗药物。
关键词
Keywords: NAMPT; TLR4; CYBB; NLRP3 inflammasome; colitis
关键词:NAMPT;TLR4;CYBB;NLRP3 炎症小体;结肠炎
1. 简介 Introduction
Inflammatory bowel diseases (IBDs) including Crohn’s disease as well as ulcerative colitis (UC) are characterized by abnormalities in intestinal homeostasis, which result in chronic inflammation [1,2]. Biological drugs (anti-TNFα therapy and/or immune modulators) are effective for treating many people with IBD. However, many patients with severe disease fail to achieve remission because of a poor responsiveness to drugs, unavailability of mechanism-specific targeted therapy, or serious adverse effects requiring treatment [3,4]. Therefore, highly sensitive and specific predictive biomarkers along with novel therapeutic strategies are required to implement effective therapeutic strategies.
包括克罗恩病和溃疡性结肠炎 (UC) 在内的炎症性肠病 (IBD) 的特征是肠道稳态异常,从而导致慢性炎症 [1,2]。生物药物(抗 TNFα 疗法和/或免疫调节剂)可有效治疗许多 IBD 患者。然而,许多重症患者由于对药物的反应不佳、机制特异性靶向治疗不可用或需要治疗的严重不良反应而未能获得缓解[3,4]。因此,需要高度敏感和特异性的预测性生物标志物以及新的治疗策略来实施有效的治疗策略。
Nicotinamide phosphoribosyltransferase (NAMPT) acts as a catalyst of the ratedetermining step of the nicotinamide adenine dinucleotide (NAD) recovery pathway and thus regulates intracellular NAD concentrations [5]. Many cell types secrete NAMPT, including monocytes and macrophages [6–8]. Accumulating evidence suggests that extracellular (e) NAMPT (visfatin) acts as a novel soluble factor with activities similar to those of cytokines, adipokines, and DAMPs [9–11]. Further, eNAMPT contributes to metabolic and inflammatory disorders such as obesity, diabetes, cancer, and particularly IBD [6,12–14]. The cellular and serum levels of eNAMPT are increased in patients with IBD who are unresponsive to anti-TNFα treatment (adalimumab or infliximab); and eNAMPT levels decrease in responsive patients to values comparable with those of healthy controls [15,16]. These findings support the conclusion that because eNAMPT levels correlate with a worse prognosis, this enzyme will serve as a potential target of drugs for effectively and safely treating IBD.
烟酰胺磷酸核糖基转移酶 (NAMPT) 作为烟酰胺腺嘌呤二核苷酸 (NAD) 恢复途径的决速步骤的催化剂,从而调节细胞内 NAD 浓度 [5]。许多细胞类型都会分泌 NAMPT,包括单核细胞和巨噬细胞 [6–8]。越来越多的证据表明,细胞外 (e) NAMPT(内脂素)作为一种新型可溶性因子,其活性类似于细胞因子、脂肪因子和 DAMP [9-11]。此外,eNAMPT 会导致代谢和炎症性疾病,例如肥胖、糖尿病、癌症,尤其是 IBD [6,12–14]。在抗 TNFα 治疗(阿达木单抗或英夫利昔单抗)无反应的 IBD 患者中,eNAMPT 的细胞和血清水平升高;和 eNAMPT 水平在有反应的患者中降低到与健康对照相当的值 [15,16]。这些发现支持以下结论:由于 eNAMPT 水平与较差的预后相关,因此该酶将作为有效和安全治疗 IBD 的潜在药物靶点。
既往关于烟酰胺磷酸核糖基转移酶(NAMPT)研究的作用机制eNAMPT 的作用主要与巨噬细胞中炎症信号的激活有关,表明 TLR4 介导的 NF-kB 信号的有效激活是通过直接结合 TLR4 发生的 [14,17–19]。此外,用 NAMPT 抑制剂或 NAD 前体改变细胞内 NAD 水平会改变 TLR4 介导的 NF-kB 激活和 NLRP3 炎性体活性,从而建立细胞内 NAD 水平与炎症之间的关系 [20]。此外,eNAMPT 可能与 NADPH 氧化酶 (NOX) 的激活相关,通过脂质筏聚集刺激 NOX 衍生的活性氧 (ROS),进而通过 TXNIP 激活 NLRP3 炎性体 [21,22]。因此,通过氧化还原信号通路激活 NLRP3 炎性体可能通过 NF-kB 信号通路(启动)和通过 NAMPT-NOX 结合的相互作用(激活)发生。
Intensive research focused on colonic-targeted drug delivery systems for topical treatment of colonic disease shows that they cause fewer systemic side effects and improve the effectiveness of oral delivery of therapeutic agents susceptible to acidic and enzymatic breakdown in the upper gastrointestinal tract [23,24]. A targeting moiety often enhances gene therapy. The inflamed colon can be targeted using a 12-residue peptide (TWYKIAFQRNRK; designated “CT” for colonic-targeting); it interacts with high affinity with the integrin α6β1, which is generated from the COOH-terminal globular domain belonging to the laminin-1 α1 chain [23,24]. Thus, the CT peptide may serve as a promising moiety for ameliorating colonic inflammation.
针对局部治疗结肠疾病的结肠靶向给药系统的深入研究表明,它们引起的全身副作用较少,并提高了口服给药对上消化道酸性和酶促分解敏感的治疗药物的有效性 [23,24]。靶向部分通常增强基因治疗。可以使用 12 残基肽(TWYKIAFQRNRK;结肠靶向指定为“CT”)靶向发炎的结肠;它与整合素 α6β1 以高亲和力相互作用,整合素 α6β1 由属于层粘连蛋白 1 α1 链的 COOH 末端球状结构域产生 [23,24]。因此,CT 肽可作为改善结肠炎症的有前途的部分。
Our present data, which were acquired from genome-wide association studies of NAMPT in humans and mouse models of colitis, identified key driver genes in macrophages associated with IBD. Notably, we found that eNAMPT interacted with TLR4 or CYBB/NOX2 in IBD tissues. Moreover, recombinant multifunctional NAMPT (rCT-NAMPT) linked to the CT peptide that harbors the essential minimal residues required for binding CYBB and TLR4, may serve as a platform for developing safe and effective treatments of IBD.
我们目前的数据是从 NAMPT 在人类和小鼠结肠炎模型中的全基因组关联研究中获得的,确定了与 IBD 相关的巨噬细胞中的关键驱动基因。值得注意的是,我们发现 eNAMPT 与 IBD 组织中的 TLR4 或 CYBB/NOX2 相互作用。此外,与 CT 肽相连的重组多功能 NAMPT (rCT-NAMPT) 具有结合 CYBB 和 TLR4 所需的基本最小残基,可作为开发安全有效的 IBD 治疗的平台。
2. 材料与方法 Materials and Methdos
2.1. 溃疡性结肠炎的公共单细胞 RNA-Seq 数据分析
选择数据集,列出样本信息使用的数据集:我们使用了两个数据集,SPC259 [25],可从人类细胞图谱中获得,以及 GSE182270 [26]。表 1 显示了数据集的简要摘要。虽然除了这两个之外还有几个数据集,例如 GSE162335 [27] 和 GSE125527 [28],但对于溃疡性结肠炎,我们选择了这两个,因为其他数据集不包含足够数量的巨噬细胞和 T 细胞,这是我们在研究中的主要关注点这项研究。由于 GSE182270 中巨噬细胞的数量有限,我们仅将其用作 DEG 和 GSE 分析的补充数据,以验证 SCP259 的主要结果。

DEG and GSEA: We performed differentially expressed gene (DEG) analysis as well as gene set enrichment analysis (GSEA) individually on the selected cells for the two datasets. Specifically, macrophage, regulatory T cells, other CD4+ T cells and cytotoxic T cells were analyzed. The latest version of Seurat package [29] was used for the DEG analysis and FGSEA package [26,29] was used for GSEA. Since the number of cells in GSE182270 is much smaller than that in SCP259, we performed the analysis as follows. (1) DEG: DEGs were identified separately for the two datasets with a p-value cutoff of 0.05. Rather than using an adjusted p-value, we used a non-adjusted one to obtain as many DEGs as possible. (2) Selection of common DEGs: We then selected those DEGs that have the same sign of log-fold change in the two datasets and then took the intersection of the two sets of DEGs, which is used for the GSEA. (3) GSEAs with common DEGs: GSEA analyses were performed with the selected DEGs with the log-fold changes for SCP259, since it might provide a better result when using a greater number of cells.
DEG 和 GSEA:我们对两个数据集的选定细胞分别进行了差异表达基因 (DEG) 分析和基因集富集分析 (GSEA)。具体而言,分析了巨噬细胞、调节性 T 细胞、其他 CD4+ T 细胞和细胞毒性 T 细胞。最新版本的 Seurat 包 [29] 用于 DEG 分析,FGSEA 包 [26,29] 用于 GSEA。由于 GSE182270 中的细胞数量远小于 SCP259 中的细胞数量,我们进行了如下分析。 (1) DEG:分别为两个数据集确定了 DEG,p 值截止值为 0.05。我们没有使用调整后的 p 值,而是使用未调整的 p 值来获得尽可能多的 DEG。 (2) 公共 DEG 的选择:然后我们选择了两个数据集中具有相同对数倍变化符号的 DEG,然后取两组 DEG 的交集,用于 GSEA。 (3) 具有共同 DEG 的 GSEA:GSEA 分析是对选定的 DEG 进行的,其中 SCP259 的对数倍变化,因为当使用更多的细胞时它可能会提供更好的结果。
2.2. 小鼠和组织培养
Wild-type C57BL/6 mice were supplied by Samtako Bio Korea (Osan, Korea). Primary bone marrow–derived macrophages (BMDMs) were harvested from mice and cultured in DMEM in the presence of M-CSF (R&D Systems, 416-ML) for 3–5 d, as described previously [30,31]. HEK293T cells (ATCC-11268; American Type Culture Collection) were cultured in in DMEM (Gibco) containing 10% FBS (Gibco), nonessential amino acids, sodium pyruvate, streptomycin (100 µg/mL) and penicillin G (100 IU/mL).
野生型 C57BL/6 小鼠由 Samtako Bio Korea (Osan, Korea) 提供。如前所述 [30,31],原代骨髓源性巨噬细胞 (BMDM) 从小鼠身上采集并在 M-CSF(R&D Systems,416-ML)存在下在 DMEM 中培养 3-5 天。 HEK293T 细胞(ATCC-11268;美国典型培养物保藏中心)在含有 10% FBS (Gibco)、非必需氨基酸、丙酮酸钠、链霉素 (100 µg/mL) 和青霉素 G (100 IU/mL) 的 DMEM (Gibco) 中培养)。
2.3. 试剂和抗体
购买试剂脂多糖(Escherichia coliO111:B4,tlrl-eblps)、三磷酸腺苷(ATP,tlrl-ATPL)、尼日利亚霉素和葡聚糖硫酸钠(DSS)购自Invivogen。
购买抗体抗NAMPT(Ab236874)的抗体购自Abcam。NAMPT(E-3)、TLR4(25)、CyBB(54.1)、CyBA(E-11)、肌动蛋白(I-19)、ASC(N-15-R)、IL-18(H-173-Y)、caspase-1 p10(M-20)、HA(12CA5)、FLAG(D-8)、GST(B-14)、Myc(9E10)、His(H-3)和V5(C-9)购自Santa Cruz Biotech。针对IkBα(L35A5)和磷酸化IkBα(14D4)的特异性抗体由细胞信号技术公司(美国马萨诸塞州丹弗斯)提供。IL-1β(AF-401-NA)和NLRP3(AG-20B-0014)分别来自R&D Systems和Adipogen。
2.4. 质粒构建
GST-NAMPT, Myc-NAMPT and Flag-TLR4 plasmids were sourced from Addgene. The full-length (FL) CYBB and mutant plasmids were previously described [32]. Plasmids encoding different regions of NAMPT (1-491, 10-116, 188-455, 456-491) were amplified by PCR from full-length NAMPT cDNA and subcloned into a pEBG derivative that codes for an N-terminal GST epitope tag flanked by the BamHI and NotI sites. Plasmids encoding different regions of TLR4 (1-811, 25-696, 717-811) were produced via PCR amplification from FL TLR4 cDNA and subcloned into a pEF derivative encoding a C-terminal Flag-tag between the BamHI and NotI sites. The pEBG-GST mammalian fusion vector and the pEF- IRES-Puro expression vector were used to create all transient constructs in mammalian cells. All constructions were sequenced using an ABI PRISM 377 automated DNA sequencer to ensure that they were 100% identical to the original sequence.
GST-NAMPT、Myc-NAMPT和Flag-TLR4质粒来源于Addgene。全长(FL)的CYBB和突变质粒以前已经描述过[32]。从NAMPT全长cDNA中扩增出编码NAMPT不同区域(1-491、10-116、188-455、456-491)的质粒,并将其亚克隆到编码BamHI和Noti位点两侧的GST N端表位标签的pEBG衍生体中。通过从FL TLR4基因中扩增出编码TLR4不同区域(1-811、25-696、717-811)的质粒,并将其亚克隆到编码BamHI和NotI位点之间的C-末端Flag-tag的PEF衍生体中。PEBG-GST哺乳动物融合载体和PEF-IRES-Puro表达载体用于在哺乳动物细胞中构建所有瞬时结构。使用ABI PRISM 377自动DNA测序仪对所有构建物进行测序,以确保它们与原始序列100%相同。
2.5. 肽类
Tat-conjugated NAMPT or NAMPT peptides were commercially produced and purified in acetate salt form to circumvent undesirable responses in the cells by Peptron (Daejeon, Republic of Korea). The endotoxin level was determined by the Limulus amebocyte lysate test (Charles River Endosafe® Endochrome-K™, R1708K, Wilmington, MA, USA) and was less than 3–5 pg/mL at the concentrations of the peptides employed in experiments.
TAT偶联的NAMPT或NAMPT多肽以醋酸盐形式商业化生产和提纯,以避免Peptron(韩国大田)在细胞中的不良反应。细菌内毒素的测定采用美国马萨诸塞州威尔明顿的Charles River Endosafe®EndoChrome-K™试剂盒(Charles River Endosafe®EndoChrome-K™,R1708K),在实验所用多肽的浓度范围内,内毒素水平小于3~5pg/mL。
2.6. 重组蛋白
To obtain recombinant rCT-NAMPT proteins, colon-targeting sequences of NAMPT amino acids (57–65), NAMPT amino acids (52–65), and CT peptides (TWYKIAFQRNRK) were cloned with an N-terminal 6xHis tag into the pRSFDuet-1 Vector (Novagen) and induced, harvested, and purified from Escherichia coli expression [30,31], in accordance with the standard protocols recommended by Novagen. The rVehicle, rCT, or rCT-NAMPT proteins were dialyzed through a permeable cellulose membrane and tested for lipopolysaccharide contamination using a Limulus amebocyte lysate assay (Bio-Whittaker) and contained <20 pg/mL at the concentrations of rVehicle, rCT, or rCT-NAMPT proteins used in the experiments.
为了获得重组的RCT-NAMPT蛋白,将NAMPT氨基酸(57-65)、NAMPT氨基酸(52-65)和CT多肽(TWYKIAFQRNRK)的结肠靶向序列克隆到pRSFDuet-1载体(Novagen)中,并按照Novagen推荐的标准程序从大肠杆菌表达[30,31]中进行诱导、收获和纯化。RVehicle、RCT或RCT-NAMPT蛋白通过可渗透的纤维素膜透析,并用凝胶变形细胞裂解试验(Bio-Whittaker)检测脂多糖污染,在实验中使用的rVehicle、RCT或RCT-NAMPT蛋白的浓度为<20pg/mL。
2.7. GST下拉、免疫印迹和免疫沉淀分析
293T and BMDMs were treated as specified and subjected to GST pull-down, Western blotting, and co-immunoprecipitation analysis, as previously reported [31–34].
293T和BMDM按规定处理,并进行GST下拉、Western blotting和免疫共沉淀分析,如先前报道的[31-34]。
293T cells were collected and lysed in NP-40 buffer supplemented with a full protease inhibitor cocktail (Roche, Basal, CH), for GST pull-down. After centrifugation, the supernatants were pre-cleared for 2 h using protein A/G beads at 4 ◦C. Pre-cleared lysates were combined with a 50% slurry of glutathione-conjugated Sepharose beads (Amersham Biosciences, Amersham, UK), and the binding reaction was incubated at 4 ◦C for 4 h. The precipitates were thoroughly rinsed with lysis buffer. Proteins bound to glutathione beads were eluted by boiling for 5 min in Sodium Dodecyl Sulfate (SDS) loading buffer.
收集293T细胞,裂解于NP-40缓冲液中,添加完全的蛋白酶抑制剂鸡尾酒(罗氏,Basal,CH),用于GST下拉。离心后,上清液在4℃下用蛋白A/G珠预净化2 h,预清的裂解液与50%谷胱甘肽偶联琼脂糖珠(amersham Bioscience,amersham,UK)的浆液结合,结合反应在4℃孵育4 h,沉淀用裂解缓冲液彻底冲洗。在十二烷基硫酸钠(十二烷基硫酸钠)负载缓冲液中煮沸5min洗脱结合在谷胱甘肽小球上的蛋白质。
Cells were collected and lysed in NP-40 buffer supplemented with a full protease inhibitor cocktail (Roche, Basal, CH) for immunoprecipitation. Whole-cell lysates were immunoprecipitated with the indicated antibodies after pre-clearing with protein A/G agarose beads for 1 h at 4 ◦C. In general, 1 mL of cell lysates was treated with 1–4 µg of commercial antibody for 8 to 12 h at 4 ◦C. After 6 h of incubation with protein A/G agrose beads, the immunoprecipitates were thoroughly washed with lysis buffer and eluted with SDS loading buffer by boiling for 5 min
收集细胞并裂解于NP-40缓冲液中,添加完全的蛋白酶抑制剂鸡尾酒(Roche,Basal,CH)进行免疫沉淀。全细胞裂解产物在蛋白A/G琼脂糖珠4◦C预澄清1h后与所示抗体进行免疫共沉淀。一般情况下,每毫升细胞裂解产物在4◦C下用1-4微克商品抗体处理8~12h。在与蛋白A/G琼脂糖珠孵育6h后,免疫沉淀物用裂解缓冲液彻底洗涤,并用十二烷基硫酸钠负载缓冲液沸腾洗脱5min。
Polypeptides were separated by SDS–polyacrylamide gel electrophoresis and then transferred to a PVDF membrane for immunoblotting (IB) (Bio-Rad, Hercules, CA, USA). Achieving immunodetection required specific antibodies. Chemiluminescence (ECL; Millipore, Burlington, MA, USA) was used to visualize an antibody bound to a target, and a Vilber chemiluminescence analyzer was used to detect it (Fusion SL 3; Vilber Lourmat, Collégien, France).
用SDS-聚丙烯酰胺凝胶电泳法分离多肽,然后转移到PVDF膜上进行免疫印迹(IB)(Bio-Rad,Hercules,CA,USA)。实现免疫检测需要特定的抗体。用化学发光法(ECL;美国马萨诸塞州伯灵顿米利波尔)显示与目标结合的抗体,并使用维尔伯化学发光分析仪进行检测(Fusion SL 3;Vilber Lourmat,Collégien,法国)。
2.8. 流式细胞术
Flow cytometry was used to quantify intracellular ROS levels in cells cultivated in serum-free media and loaded with the redox-sensitive dyes 2 µM dihydroethidium (DHE for O2−; Calbiochem) or 1 µM 20,70-dichlorodihydrofluorescein diacetate (H2DCFDA for H2O2; Calbiochem) [32]. mAbs were incubated at 4 ◦C for 20–30 min to determine cell surface protein expression, and cells were fixed using Cytofix/Cytoperm Solution (BD Biosciences) and, in certain cases, followed by mAb incubation to detect intracellular proteins. [35]. The mAb clones that were used were the following: ITGA6 (GoH3, BD Pharmingen™) and ITGB1 (HM β1-1, BD Pharmingen™). The cells were washed completely and quickly with pulse spinning before being analyzed in a FACSCalibur (BD Biosciences, San Jose, CA, USA). CellQuest software (BD Biosciences) was used to visualize the data, and FlowJo software was used to analyze it (Tree Star, Ash-land, OR, USA).
用流式细胞术对无血清培养的细胞内ROS水平进行定量,并加载氧化还原敏感染料2微米二氢乙啶(DHE)(O2−;钙化)或1微米20,70-二氯二氢荧光素二乙酸酯(H2DCFDA用于过氧化氢;钙化)[32]。单抗在4◦C下孵育20-30分钟以检测细胞表面蛋白的表达,细胞用细胞固定液(BD Biosciences)固定,在某些情况下,随后单抗孵育以检测细胞内蛋白。[35][font=宋体]。所用的单抗克隆如下:ITGA6(GoH3,BD Pharmingen™)和ITGB1(HMβ1-1,BD Pharmingen™)。在FACSCalibur(BD Biosciences,San Jose,CA,USA)中分析之前,用脉冲旋转完全且快速地洗涤细胞。使用CellQuest软件(BD Biosciences)进行数据可视化,FlowJo软件进行数据分析(Tree Star,Ash-land,OR,USA)。
2.9. 酶联免疫吸附试验
TNF-α, IL-6, IL-1β and IL-18 levels were measured in cell culture supernatants and mouse serum using the BD OptEIA ELISA system (BD Pharmingen). All assays were carried out exactly as the manufacturer instructed.
用BD OptEIA ELISA系统(BD Pharmingen)检测细胞培养上清液和小鼠血清中肿瘤坏死因子α、IL 6、IL 1β和IL 18水平。所有的化验都严格按照制造商的指示进行。
2.10. 慢病毒shRNA生产
For silencing murine ITGA6 and ITGB1 in primary cells, pLKO.1-based lentiviral CaMKKb shRNA constructs (sc-38952-SH) and LKB1 shRNA constructs (sc-35817-SH) were obtained from Santa Cruz Biotechnology. GIPZ Lentiviral Mouse Itga6 shRNA constructs (RMM4431-200328849, RMM4431-200410349 and RMM4431-200411464) and GIPZ Lentiviral Mouse Itgb1 shRNA constructs (RMM4431-200340920, RMM4431-200350229, RMM4431200386104 and RMM4431-200400706) were procured from Open Biosystems. Lentiviruses were generated via transient transfection utilizing packaging plasmids (pMDLg/pRRE, pRSV-Rev, and pMD2.VSV-G, sourced from Addgene) after Lipofectamine 2000-mediated transient transfection into HEK293T cells, as reported previously. At 72 h after transfection, the virus-containing medium was collected and concentrated by ultracentrifugation. Lentiviral vector titration was calculated using 293T cells, and the resulting lentiviruses were transduced into BMDMs, as previously described [32].
为了在原代细胞中沉默小鼠ITGA6和ITGB1,从Santa Cruz生物技术公司获得了基于pLKO.1的慢病毒CaMKKbshRNA构建体(sc-38952-SH)和LKB1shRNA构建体(sc-35817-SH)。GIPZ慢病毒小鼠Itga6shRNA构建体(RMM4431-200328849、RMM4431-200410349和RMM4431-200411464)和GIPZ慢病毒小鼠Itgb1shRNA构建体(RMM4431-200340920、RMM4431-200350229、RMM4431200386104和RMM4431-200400706)购自Open BiosSystems。根据先前的报道,慢病毒是在脂质体2000介导的瞬时转染HEK293T细胞后,利用包装载体(pMDLg/pRRE、pRSV-Rev和pMD2.VSV-G,来源于Addgene)瞬时转染产生的。72h后收集含病毒培养液,超速离心法浓缩。使用293T细胞计算慢病毒载体滴定,并将产生的慢病毒转导入BMDM,如前所述[32]。
2.11. 慢病毒在体内的转导
As previously stated, concentrated lentiviral particles were frozen at 4 ◦C and diluted in PBS and polybrene (8 g/mL final concentration; Sigma) to give a dose of 1 × 10 [10] pfu in a 100 L injection volume [32]. Mice were intravenously injected with a lentivirus expressing nonspecific shRNA (shNS) or shRNA specific for ITGA6 (sh ITGA6) or ITGB1 (shITGB1) 2 times and then orally administered DSS (Acute or chronic Colitis) and rCT-NAMPT; then, desired experiments were performed.
如前所述,浓缩的慢病毒颗粒在4◦C下冷冻,并在PBS和聚丙稀(最终浓度为8g/mLSigma)中稀释,在100L注射量中得到1×10[10]pfu的剂量[32]。小鼠静脉注射表达非特异性shRNA(ShNS)或针对ITGA6或ITGB1的shRNA(ShITGA6)或ITGB1(ShITGB1)的慢病毒2次,然后口服DSS(急性或慢性结肠炎)和RCT-NAMPT,然后进行所需的实验。
2.12. 小鼠结肠炎模型的建立
As previously described, DSS-induced acute or chronic colitis mouse models were developed using 6-week-old C57BL/6 female mice (Samtako, Osan, Republic of Korea) [36]. To assess the trigger of acute colitis, mice were treated with 3% (w/v) dextran sodium sulfate (molecular weight: 36,000–50,000 kDa, MP Biomedicals, Santa Ana, CA, USA) dissolved in drinking water that was given ad libitum. An acute colitis model was transduced with Lenti-shNS or Lenti-shITG virus (1 × 1011 pfu/kg) on days -7 and -14 via i.v. before DSS treatment. While the mice were treated with 3% DSS for 6 days, rVehicle, rCT or rCT-NAMPT (50 µg/kg) was i.p. injected 8 times. The survival of mice was tracked for 12 days; mortality was recorded for n = 15 mice per group. The survival of the mice model transduced with Lenti-shNS had similar effects to the WT control. A chronic colitis model was treated with 3% DSS for 7 days at 3 cycles and water for 14 days in the interval of the cycle. The rVehicle, rCT or rCT-NAMPT (50 µg/kg) was i.p. injected at 2 cycles together with DSS treatment. The survival of mice was followed for 9 weeks; mortality was recorded for n = 15 mice per group. To calculate causality, account for perturbations, and reduce bias, a randomization method was used to randomly assign mice to either a treatment group or a control group (or multiple intervention groups). The DSS solutions were freshly prepared every two days. The non-DSS-fed mice in the control group had access to sterile distilled water. The humane endpoint for body weight loss (euthanasia required) is 20% (as compared to the original body weight of an animal). Without an approved exception request, body weight loss could not exceed 20%.
如前所述,DSS诱导的急性或慢性结肠炎小鼠模型是使用6周大的C57BL/6雌性小鼠(韩国Samtako,Osan)[36]建立的。为了评估急性结肠炎的触发因素,小鼠被用3%(w/v)葡聚糖硫酸钠(分子量:36,000-50,000 kDa,MP Biomedals,Santa Ana,CA,USA)溶解在随意给予的饮用水中治疗。用Lenti-shNS或Lenti-shITG病毒(1×1011pfu/kg)分别于-7天和-14天静脉注射建立小鼠急性结肠炎模型。在DSS治疗前。3%DSS灌胃6d后,rVehicle、RCT或RCT-NAMPT(50µg/kg)腹腔注射。注射了8次。追踪小鼠存活12天,记录每组15只小鼠的死亡率。Lenti-shNS转导小鼠模型的存活效果与WT对照组相似。慢性结肠炎模型用3%DSS治疗7天,共3个周期,用水治疗14天,间隔3个周期。RVehicle、RCT或RCT-NAMPT(50微克/公斤)ip。在DSS治疗的同时,注射2个周期。小鼠存活9周,每组记录15只小鼠的死亡率。为了计算因果关系、解释扰动和减少偏差,使用随机化方法将小鼠随机分配到治疗组或对照组(或多个干预组)。DSS溶液每两天新鲜配制一次。对照组中未喂食DSS的小鼠饮用无菌蒸馏水。体重减轻(需要安乐死)的人道终点是20%(与动物最初的体重相比)。没有批准的例外请求,体重下降不能超过20%。
2.13. 临床评分与组织学
Body weight, occult or gross blood loss per rectum, and stool consistency were measured every other day during the colitis induction to obtain the clinical score. The clinical score was determined by two trained investigators who were not aware of the treatment. Mouse distal colon tissues were fixed in 10% formalin and embedded in paraffin for immunohistochemistry. Four-millimeter paraffin slices were cut and stained with hematoxylin and eosin (H&E). As previously mentioned, a board-certified pathologist (Dr. Min-Kyung Kim, Kim Min-Kyung Pathology Clinic, Seoul, Korea) independently scored each organ segment without prior knowledge of the therapy groups [36].
在结肠炎诱导期间每隔一天测量体重、每条直肠的隐匿性或大便失血量以及大便稠度,以获得临床评分。临床评分是由两名训练有素的调查人员决定的,他们并不知道这种治疗。小鼠远端结肠组织用10%福尔马林固定,石蜡包埋,免疫组织化学染色。切取4 mm厚石蜡切片,苏木精伊红(H&E)染色。如前所述,委员会认证的病理学家(韩国首尔Kim Min-Kyung病理诊所的Kim Min-Kyung博士)在事先不知道治疗组的情况下对每个器官片段进行独立评分[36]。
2.14. 统计分析
All data are reported as mean ± SD and were analyzed using the Student’s t-test with a Bonferroni adjustment or ANOVA for multiple comparisons. The statistical software program SPSS (Version 12.0) was used to conduct the analyses (SPSS, Chicago, IL, USA). At p 0.05, differences were judged to be significant. Data for survival were graphed and analyzed using the Kaplan–Meier product limit method, with the log-rank (Man-tele-Cox) test for comparisons in GraphPad Prism (version 5.0, La Jolla, CA, USA).
所有数据均以均值±标准差的形式报告,并使用带Bonferroni调整的学生t检验或ANOVA进行多次比较。使用统计软件程序SPSS(12.0版)进行分析(美国伊利诺伊州芝加哥SPSS)。在p0.05处,差异被判断为显著。生存数据用Kaplan-Meier乘积极限方法绘制和分析,用对数等级(Man-tele-Cox)检验在GraphPad Prism(版本5.0,La Jolla,CA,USA)中进行比较。
3. 结果 Result
3.1. ENAMPT和CyBB过表达可能导致IBD慢性炎症
Analysis of the datasets SCP259 and GSE182270 revealed that overexpressed NAMPT and CYBB (the latter also known as NOX2 and gp91Phox) may cause chronic activation of the NLRP3 inflammasome in UC (Figure 1A and Figure S1). The gastrointestinal macrophage participates in chronic inflammation in IBD, and we therefore used common macrophagespecific DEGs to check the pathways, utilizing the pathway view tool [37]. The NAMPT– CYBB interaction is associated with downstream inflammatory signaling [22]. eNAMPT stimulates the NOX-mediated redox regulatory pathway, and NAMPT associates with TLR4 [11,17,18], possibly leading to upregulation of NLRP3 and pro-IL1β expression via the NF-κB signaling pathway (Figure 1A). Overexpression of eNAMPT triggers the M1-skewed transcriptional program in macrophages [22]. These and our findings on the overexpression of NAMPT and CYBB in inflamed tissues of patients with UC led us to analyze single-cell RNA-seq data in more detail, focusing on the pathways associated with activation of the NLRP3 inflammasome through the interaction of eNAMPT with NOXs.
对数据集SCP259和GSE182270的分析表明,NAMPT和CyBB(后者也称为NOX2和gp91Phox)的过度表达可能导致UC中NLRP3炎症体的慢性激活(图1A和图S1)。胃肠巨噬细胞参与IBD的慢性炎症,因此我们使用普通的巨噬细胞特异性DEG来检查途径,利用途径查看工具[37]。NAMPT-CyBB相互作用与下游炎症信号有关[22]。ENAMPT刺激NOX介导的氧化还原调节途径,并与TLR4[11,17,18]结合,可能通过NF-βB信号通路上调NLRP3和前-IL1κ的表达(图1A)。ENAMPT的过度表达在巨噬细胞中触发了M1扭曲的转录程序[22]。这些以及我们对UC患者炎症组织中NAMPT和CYBB过表达的发现促使我们更详细地分析了单细胞RNA-SEQ数据,重点关注了通过eNAMPT与NOxs相互作用激活NLRP3炎症小体的相关途径。
图1:ENAMPT在溃疡性结肠炎(A)NAMPT与TLR4或NADPH氧化酶介导的NLRP3炎症小体激活的关系中起着关键作用。(B)SCP259中NLRP3炎症体激活前后巨噬细胞关键基因表达的小提琴图谱。(C)GSE182270中NLRP3炎症体激活周围关键基因表达的小提琴图谱。


图S1:巨噬细胞炎症与健康DEG分析结果的路径图。(A)具有来自SCP259和GSE182270的共同DEGS,(B)仅具有来自SCP259的DEGS。有了共同的DEGS,我们可以识别(1)可能由内脂素(NAMPT)-GP91(CybB)相互作用激活的NLRP3炎症体;(2)NF-κB信号通路。在(B)中,两条信令通路的激活更为清楚。
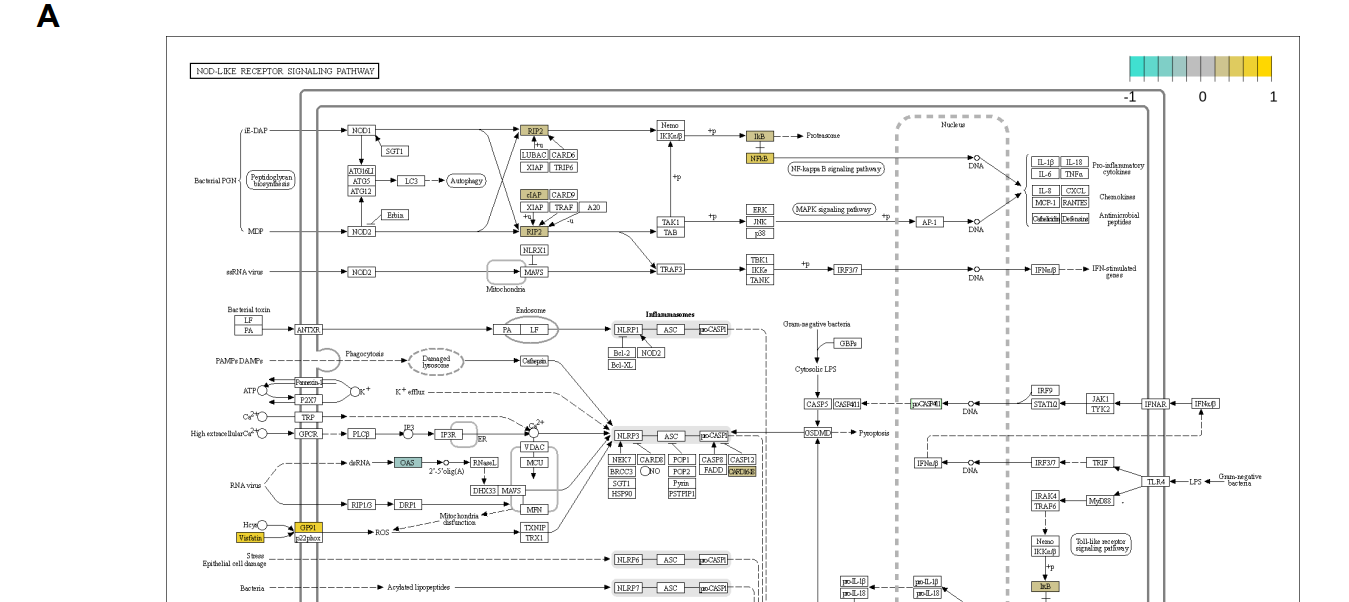
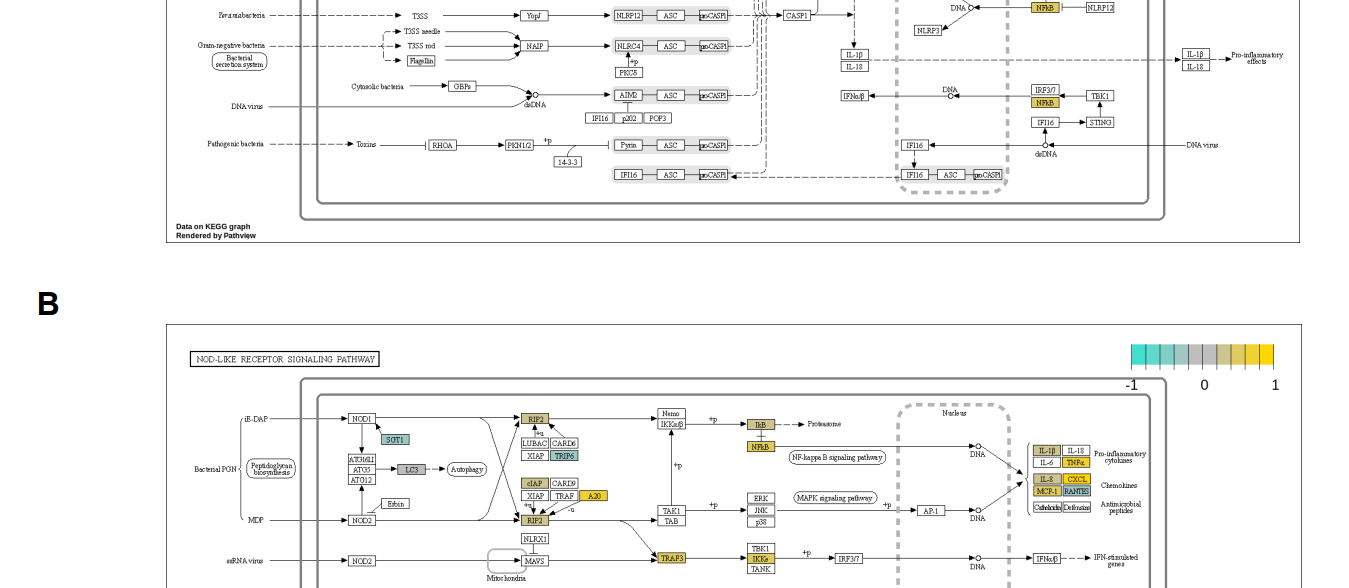
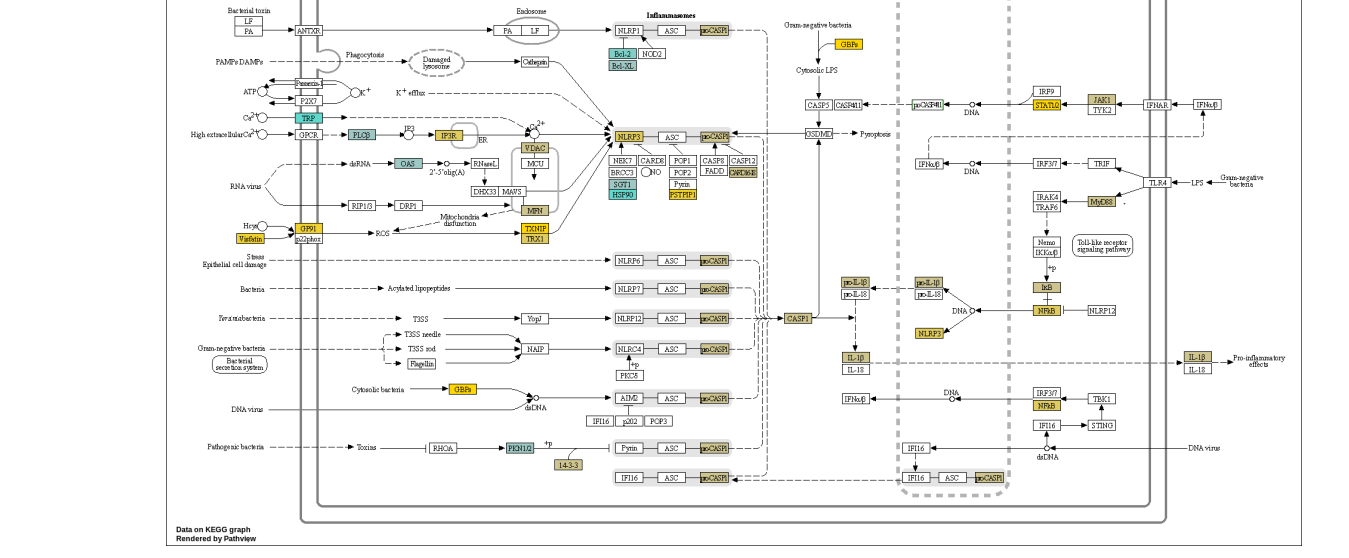

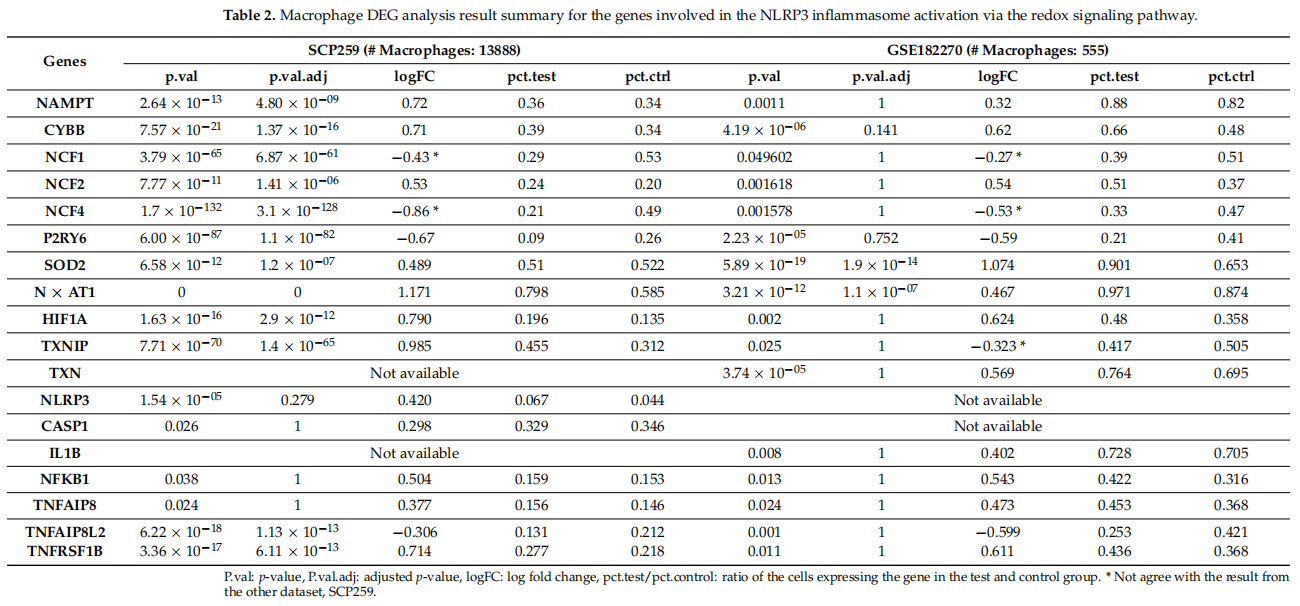
Table 2 shows the DEGs associated with NLRP3 inflammasome activation via the redox signaling pathway (activation), with focus on the NAMPT–NOX association and NF-κB signaling (priming). Violin plots (Figure 1B,C) show the expression of these genes in macrophages (Figure 1A). An increase in ROS via the NAMPT–NOX interaction is evident, because ROS-related genes such as SOD2, NEAT1, and HIF1A are overexpressed in inflamed UC samples (p < 0.05). The percentages of macrophages expressing NAMPT were similar in inflamed and control tissues, and their expression levels were higher in inflamed vs. control samples. Although the adjusted p-value for the fold-change in NAMPT expression in GSE182270 was not statistically significant, this may be explained by the relatively small number of macrophages.
表2显示了与NLRP3炎症体激活相关的DEGS,这些DEGS通过氧化还原信号通路(激活),重点是NAMPT-NOX结合和NF-κB信号(启动)。小提琴曲线图(图1B,C)显示了这些基因在巨噬细胞中的表达(图1A)。通过NAMPT-NOX相互作用,ROS明显增加,因为ROS相关基因,如SOD2,NEAT1,和HIF1a在炎症UC样本中过度表达(p<0.05)。炎症组织和对照组巨噬细胞表达NAMPT的百分率相似,而炎症组织中NAMPT的表达水平高于对照组。虽然GSE182270中NAMPT表达倍数变化的调整p值没有统计学意义,但这可能是由于巨噬细胞数量相对较少所致。
The gene set enrichment results (Table S1) show that in macrophages, many of the immune signals are upregulated in UC compared with those in the healthy colon. These signals include interleukin (IL)s, the innate immune system, Toll-like receptor cascades, TNFα signaling via NF-κB, inflammatory responses, cytokine signaling in the immune system, and IL-4 and IL-13 signaling (Figure 1A). These signaling events are involved in NLRP3 activation and in priming signaling pathways (Figure 1A). Further, genes associated with IFNγ signaling, antigen processing and presentation, as well as MHC Class II antigen presentation, were downregulated. Taken together, these data indicate that eNAMPT contributes to UC and that macrophages in UC transmit higher levels of innate immune signals associated with NAMPT–CYBB-driven signaling pathways.
基因集浓缩结果(表S1)显示,在巨噬细胞中,与健康结肠相比,UC中的许多免疫信号上调。这些信号包括白介素s、先天免疫系统、Toll样受体级联、肿瘤坏死因子α信号通过核因子-κB、炎症反应、免疫系统中的细胞因子信号以及IL-4和IL-13信号(图1A)。这些信号事件参与了NLRP3的激活和启动信号通路(图1A)。此外,与干扰素γ信号、抗原处理和呈递以及MHC第二类抗原呈递相关的基因下调。综上所述,这些数据表明eNAMPT在UC中起作用,UC中的巨噬细胞传递与NAMPT-CyBB驱动的信号通路相关的更高水平的先天免疫信号。
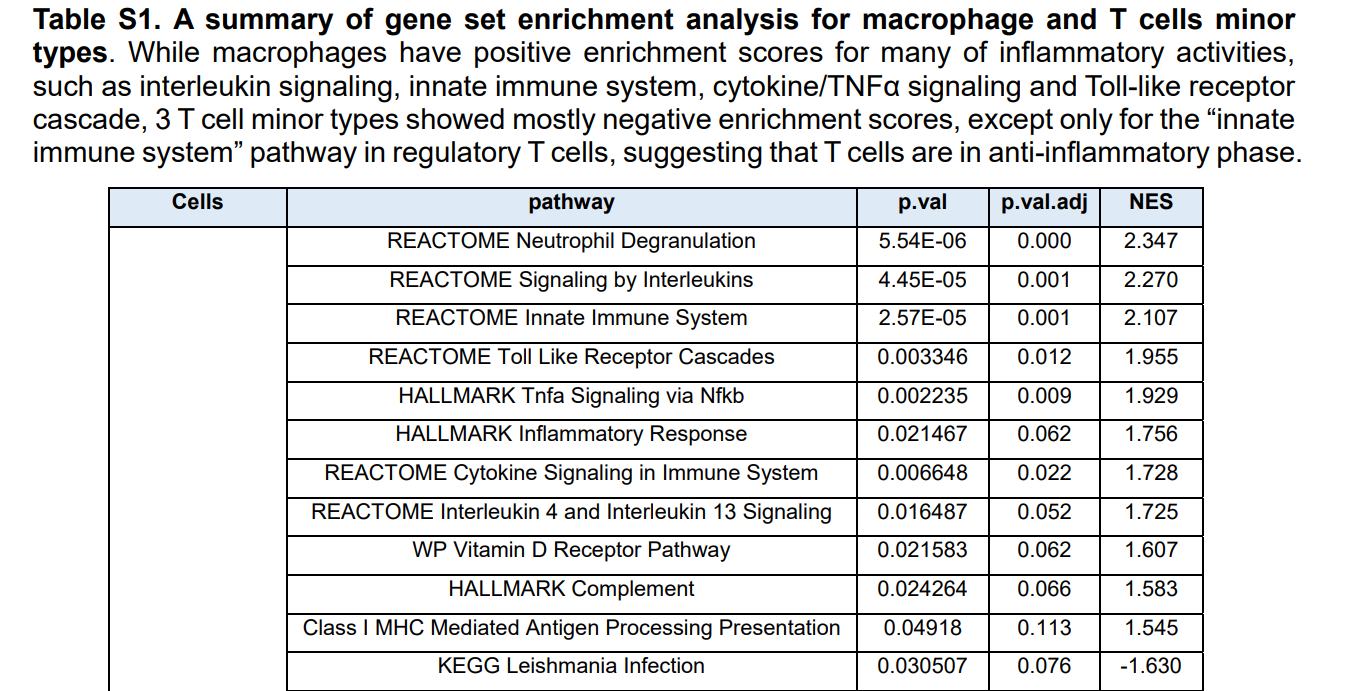
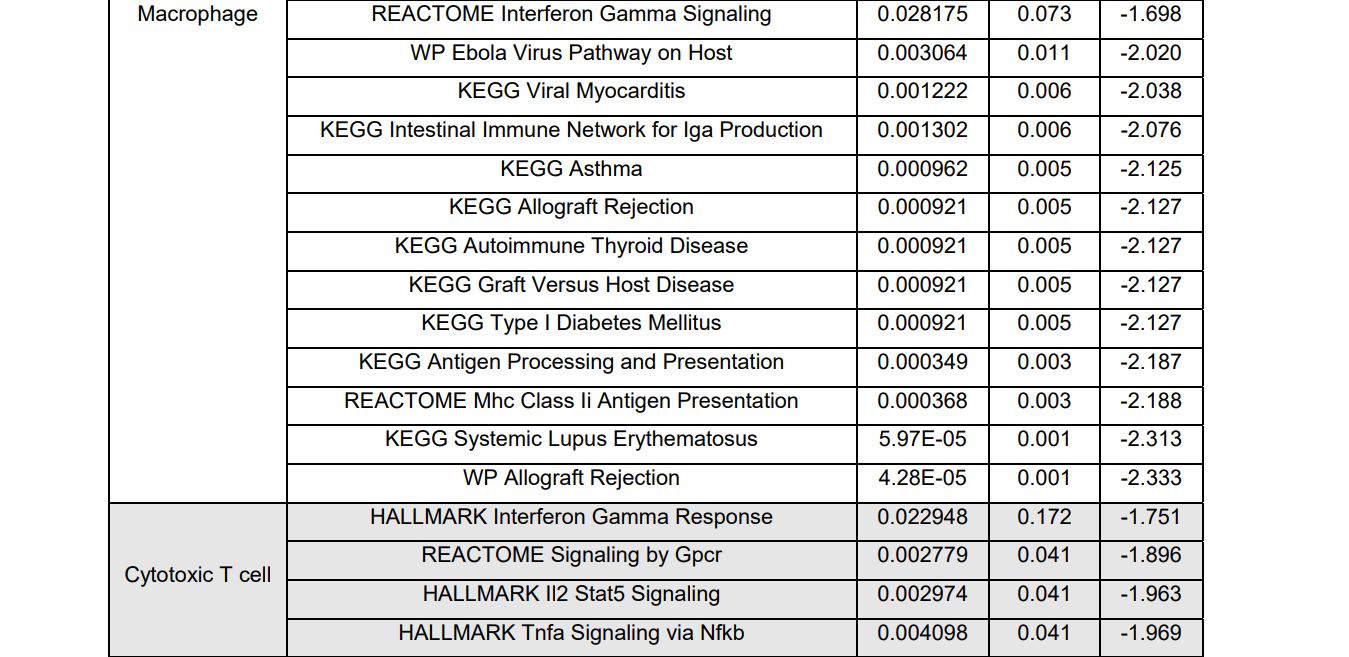
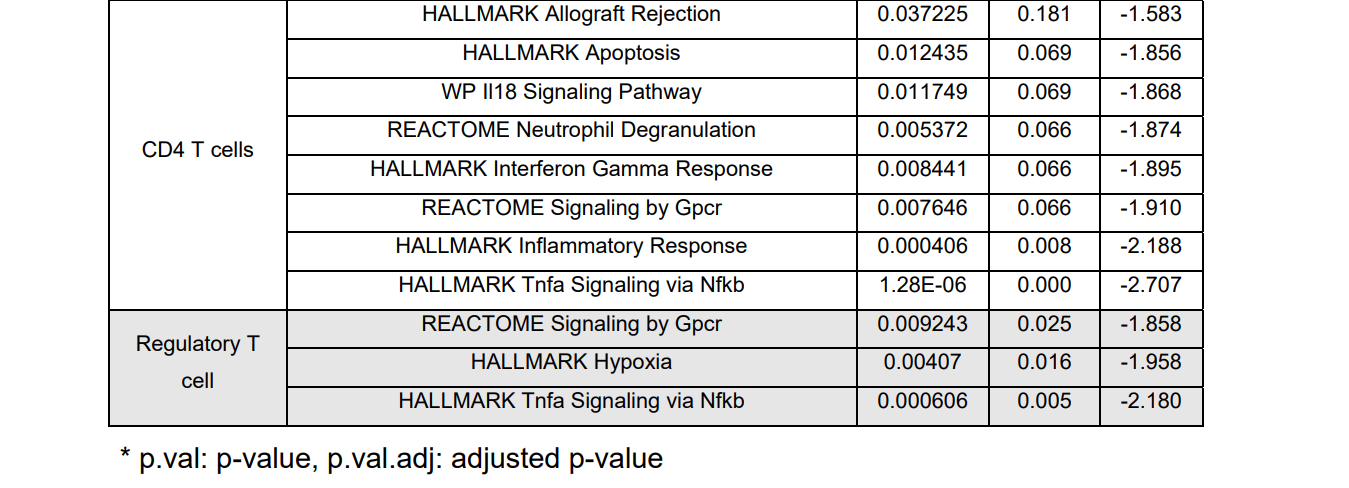
表S1:巨噬细胞和T细胞亚型基因集浓缩分析综述。巨噬细胞的许多炎症活动,如白细胞介素性信号、先天免疫系统、细胞因子/肿瘤坏死因子α信号和Toll样受体级联等均有正的富集值,但除调节性T细胞中的“先天免疫系统”途径外,3种T细胞亚型的富集率大多为负值,提示T细胞处于抗炎阶段。
3.2. ENAMPT与CyBB和TLR4相互作用
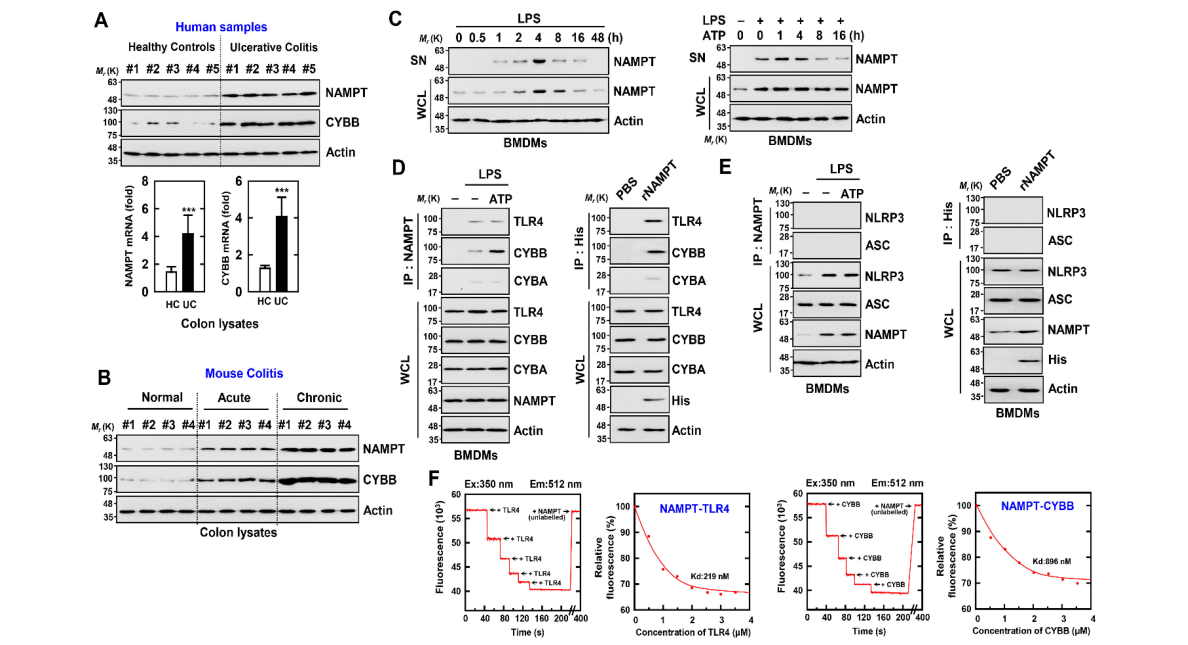
We further analyzed NAMPT and CYBB expression in colon tissue sections from normal subjects and patients with UC. The mRNA and protein levels of NAMPT and CYBB were approximately three to four times higher in the colon tissues of patients with UC compared with those of controls (Figure 2A). Further, the expression of NAMPT and CYBB markedly increased in association with increased disease severity in normal mice or those with acute or chronic colitis (Figure 2B).
我们进一步分析了正常受试者和UC患者结肠组织切片中NAMPT和CYBB的表达。UC患者结肠组织中NAMPT和CyBB的mRNA和蛋白水平约为对照组的三到四倍(图2A)。此外,在正常小鼠或急性或慢性结肠炎小鼠中,随着疾病严重程度的增加,NAMPT和CyBB的表达显著增加(图2B)。
图2:eNAMPT直接与α或TLR4结合(A)正常人和UC患者被用于IB与αNAMPT和TLR4结合。全细胞裂解液(WCL)用于IB,α肌动蛋白(上)。图中显示了来自五个独立的健康对照和患者的代表性凝胶图像。实时定量聚合酶链式反应检测NAMPT和CYBB基因的表达(下)。图中显示了10个正常人和UC患者的5个数据。统计学意义通过学生t检验结合Bonferroni校正(*p<0.001)与人类正常进行评估。(2)对正常小鼠、急性结肠炎小鼠和慢性结肠炎小鼠的结肠进行α-NAMPT和α-CyBB的IB分析。使用WCL和α肌动蛋白作为IB。每种条件下进行生物重复试验(n=10)。(C)BMDM先用内毒素(100 ng/m L)激活,持续时间(左),然后用内毒素(100 ng/m L)预刺激4h,再用1 mM三磷酸腺苷(1 MM)刺激(右)。α-NAMPT和α-NAMPT或α-肌动蛋白联合培养上清液中的IB。(d,E)BMDM用脂多糖预处理,用ATP刺激(左)或与rNAMPT(1µg/mL)孵育2 h(右)。BMDM用αNAMPT或αHis进行IP治疗,IB用αTLR4、αCyBB或αCyba(D),IB用αAsc或αNLRP3(E)。用α-TLR4、α-CYBB、α-CYBA、α-ASC、α-NLRP3或α-肌动蛋白作为IB。(F)用未标记的NAMPT滴定荧光标记的TLR4或CyBB,用曲线拟合分析确定Kd(219和896 nm)。这些数据来自五个独立的实验,这些实验产生了类似的结果。

To confirm a role for eNAMPT in activating the NLRP3 inflammasome in macrophages, we examined whether the interaction of eNAMPT with TLR4 or CYBB was involved. A two-signal model was proposed to explain activation of the inflammasome by NLRP3 [22]. Signal 1 is a priming signal created by microbial components or endogenous cytokines that mediates NF-κB activation and subsequent increase of NLRP3 and prointerleukin-1β levels. Various molecules, including extracellular ATP and pore-forming toxins, transmit inflammatory activation signals (signal 2). Several molecular or cellular events activate the NLRP3 inflammasome, such as ion flux, mitochondrial dysfunction, ROS generation, and lysosomal damage [22 ,38].
为了证实eNAMPT在激活巨噬细胞中NLRP3炎症体中的作用,我们检测了eNAMPT是否与TLR4或CyBB相互作用。提出了一个双信号模型来解释NLRP3[22]对炎症体的激活。信号1是由微生物成分或内源性细胞因子产生的启动信号,它介导了NF-κB的激活以及随后NLRP3和前白介素1β水平的增加。各种分子,包括细胞外三磷酸腺苷和致孔毒素,传递炎症激活信号(信号2)。一些分子或细胞事件激活NLRP3炎症体,如离子通量、线粒体功能障碍、ROS生成和溶酶体损伤[22,38]。
First, we found that NAMPT expressed by TLR4 (LPS) or an NLRP3 inflammasome inducer (LPS/ATP) increased intracellular (i) NAMPT expression and markedly increased extracellular (e) NAMPT expression in macrophages (Figure 2C). Further, eNAMPT interacted with TLR4 or CYBB in the presence of LPS; and eNAMPT treated with recombinant NAMPT protein interacted with TLR4 or CYBB in macrophages. In contrast, the NAMPT interaction with CYBA was negligible in macrophages (Figure 2D). Further, an interaction between NAMPT and NLRP3 or ASC was undetectable (Figure 2E). Furthermore, the in vitro interaction between NAMPT and TLR4 or CYBB, as evaluated by a fluorescence binding experiment with recombinant proteins and fluorescently labeled TLR4 or CYBB with NAMPT, revealed a sufficiently high affinity (TLR4, 219 nM; CYBB, 896nM) (Figure 1). Together, the data suggest that the interaction of eNAMPT with TLR4 or CYBB mediates the activation of the NLRP3 inflammasome in inflammatory colitis, which indicates the potential clinical importance of these events.
首先,我们发现,由TLR4(脂多糖)或NLRP3炎性小体诱导剂(LPS/ATP)表达的NAMPT增加了巨噬细胞内(I)NAMPT的表达,并显著增加了细胞外(E)NAMPT的表达(图2C)。此外,在脂多糖存在下,eNAMPT可与TLR4或CyBB相互作用,经重组NAMPT处理的eNAMPT可与巨噬细胞中的TLR4或CyBB相互作用。相反,在巨噬细胞中,NAMPT与CyBA的相互作用可以忽略不计(图2D)。此外,NAMPT与NLRP3或ASC之间的相互作用未被检测到(图2e)。此外,NAMPT与TLR4或CyBB的体外相互作用,通过重组蛋白的荧光结合实验和用NAMPT标记TLR4或CyBB的荧光标记,显示出足够高的亲和力(TLR4,219 nM;CYBB,896 nM)(图1)。总之,这些数据表明,eNAMPT与TLR4或CyBB的相互作用介导了炎性结肠炎中NLRP3炎症体的激活,这表明这些事件具有潜在的临床重要性。
3.3. TLR4与CyBB结合所必需的NAMPT氨基酸序列
To identify the amino acid (aa) residues in NAMPT that interact with TLR4 and CYBB, and we constructed vectors that express full-length and mutant NAMPT, TLR4, and CYBB. NAMPT comprises N-terminal, middle (NAPRTase), and C-terminal domains (Figure 3A). To identify the domain required for the interaction between NAMPT and TLR4, we employed constructs tagged with GST or Myc–NAMPT and Flag–TLR4. In 293T cells, the N-terminus of NAMPT bound TLR4 and the extracellular leucine-rich region (LRR domain) of TLR4 were essential for the interaction with NAMPT (Figure 3A).
为了鉴定NAMPT中与TLR4和CyBB相互作用的氨基酸残基,我们构建了表达全长和突变型NAMPT、TLR4和CyBB的载体。NAMPT由N-末端、中间(NAPRTase)和C-末端结构域组成(图3A)。为了确定NAMPT和TLR4之间相互作用所需的结构域,我们使用了标记有GST或Myc-NAMPT和Flag-TLR4的结构。在293T细胞中,NAMPT结合的TLR4的N端和TLR4的胞外富含亮氨酸的区域(LRR结构域)是与NAMPT相互作用所必需的(图3A)。
The cell-penetrating TAT peptide (GRKKRRQRRRPQ) overcomes the lipophilic barrier of cellular membranes and thus delivers large molecules as well as small particles into the cell where they exert their activities [39]. To investigate in detailed the sequence of the N-terminus of NAMPT, we constructed TAT–NAMPT peptides (separated by 20 aa) included in the stretch aa 10-116 of NAMPT. We incubated the TAT–NAMPT peptides with Myc–NAMPT and Flag–TLR4 expressed in 293T cells and immunoprecipitated (IP) the complexes with a Flag antibody. Treatment with TAT–NAMPT (aa 40–69) diminished the binding of NAMPT to TLR4, indicating that this region binds TLR4 (Figure 3B left). To determine the minimum aa sequence necessary for NAMPT–TLR4 binding, we utilized the PredictProtein software (https://predictprotein.org; accessed on 9 July 2022), which predicts elements of protein function and structure using database searches, homologybased inference, machine learning, and artificial intelligence. [40]. PredictProtein predicted that aa 57–65 of NAMPT were required for binding TLR4, consistent with findings that NAMPT–TLR4 binding in 293T cells was diminished by TAT–NAMPT (aa 57–65) in a concentration-dependent manner (Figure 3B right).
穿透细胞的TAT肽(GRKKRRQRRRPQ)克服了细胞膜的亲脂屏障,从而将大分子和小颗粒输送到细胞中,在细胞中发挥其活性[39]。为了详细研究NAMPT的N-末端序列,我们构建了包含在NAMPT的延伸AA10-116中的TAT-NAMPT多肽(间隔20个氨基酸)。我们将TAT-NAMPT多肽与Myc-NAMPT和Flag-TLR4在293T细胞中表达,并将其与Flag抗体进行免疫沉淀(IP)。用TAT-NAMPT(AA 40-69)处理后,NAMPT与TLR4的结合减弱,表明该区域与TLR4结合(图3B左)。为了确定NAMPT-TLR4结合所需的最小AA序列,我们利用了PredidicProtein软件(https://predictprotein.org;于2022年7月9日访问),该软件通过数据库搜索、基于同源的推理、机器学习和人工智能来预测蛋白质功能和结构的元件。[40][老外谈]。PredidicProtein预测,与TLR4结合需要NAMPT的AA 57-65,这与TAT-NAMPT(AA 57-65)以浓度依赖的方式减少293T细胞中NAMPT-TLR4结合的发现一致(图3B右侧)。
Next, we investigated the region of NAMPT that bound CYBB in 293T cells. The N-terminus of NAMPT bound CYBB, and extracellular domain 2 of CYBB was required for its interaction with NAMPT (Figure 3C). Further, aa 40–69 of NAMPT, the TLR4 binding residues, bound CYBB (Figure 3D left). PredictProtein predicted that NAMPT aa 52–56 as those that that bind CYBB, consistent with the decrease in NAMPT–CYBB binding in 293T cells co-expressing TAT–NAMPT (aa 52–56) in a concentration-dependent manner (Figure 3D right).
接下来,我们研究了293T细胞中与CyBB结合的NAMPT区域。NAMPT的N端与CyBB结合,其与NAMPT的相互作用需要CyBB的胞外区2(图3C)。此外,NAMPT的AA 40-69是TLR4结合残基,与CyBB结合(图3D左)。PredicatProtein预测NAMPT AA 52-56与CYBB结合的那些,与293T细胞中NAMPT-CyBB结合的减少一致,以浓度依赖的方式共表达TAT-NAMPT(AA 52-56)(图3D右)。
图3:NAMPT(AA 57-65和AA 52-65)是与TLR4和CyBB结合的重要区域。(A)NAMPT和TLR4(上)结构示意图。将GST-NAMPT及其突变体和FLAG-TLR4分别导入293T细胞。对293T细胞进行谷胱甘肽转移酶(GST)下拉,然后用α标记的IB。使用WCL与αGST、αFLAG或α肌动蛋白(左下)一起用于IB。将Flag-TLR4及其突变体和Myc-NAMPT分别导入293T细胞。293T细胞用于αMyc的IP,其次是IB的α标记。使用WCL与αMyc、αFLAG或α肌动蛋白(下,右)一起用于IB。(B)将Myc-NAMPT和Flag-TLR4分别导入293T细胞,处理NAMPT多肽或其突变体6h(左侧5µM,右侧1、5、10µM)。293T细胞用于α标记的IP,IB用于αMyc。使用WCL与αMyc、αFLAG或α肌动蛋白联合用于IB。(C)环卫总站结构示意图(上)。将GST-NAMPT或其突变体和V5-CyBB(左下)分别导入293T细胞。将GST-CyBB或其突变体和Myc-NAMPT(下,右)分别导入293T细胞。对293T细胞进行谷胱甘肽转移酶下拉,然后用αV5或αMyc进行IB。用α-GST、α-MYC、α-V5或α-肌动蛋白作为IB。(D)将Myc-NAMPT和V5-CyBB分别导入293T细胞,处理NAMPT多肽或其突变体6h(左侧5µM,右侧1、5、10µM)。293T细胞用于αV5的IP,IB用于αMyc。使用WCL与αMyc、αFLAG或α肌动蛋白联合用于IB。(E)将BMDM与rNAMPT(1、5、10微克/毫升)孵育2小时。BMDM用αHis进行IP,然后用αTLR4、αCybB或αCyba进行IB。用α-TLR4、α-CYBB或α-CYBA、α-His或α-肌动蛋白作为IB。这些数据来自七个独立的实验,它们产生了可比的结果(A-E)。REF STAR:具有结合伙伴的结合结构域。
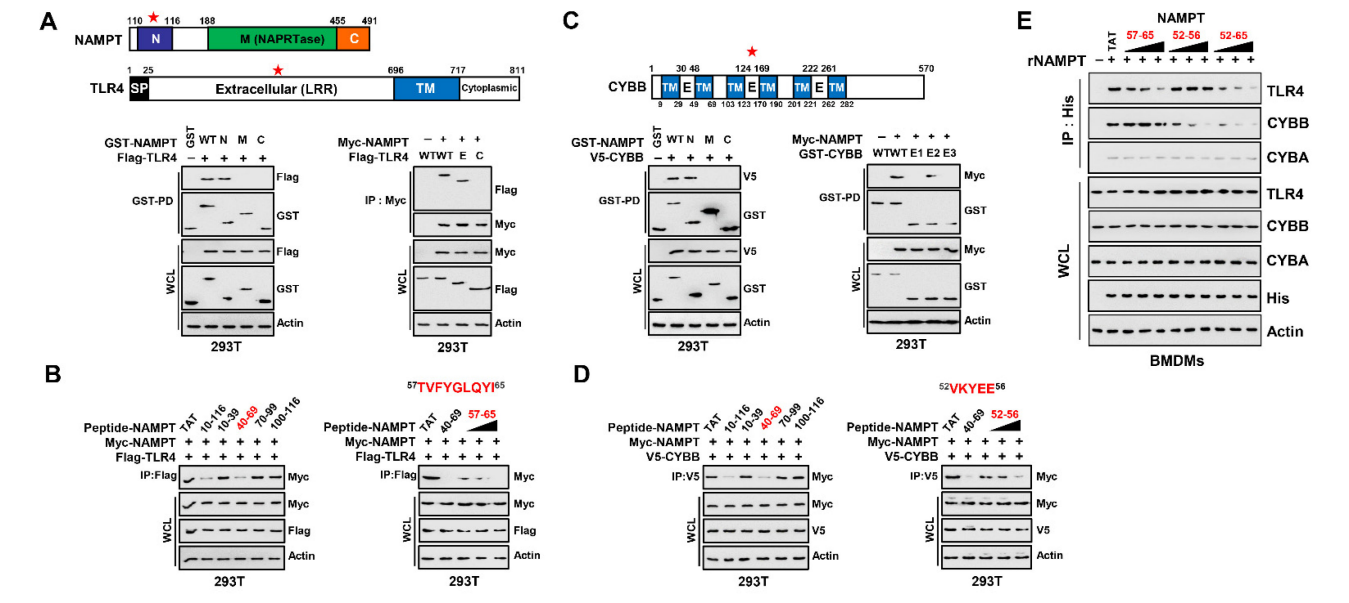

We next tested whether TAT–NAMPT aa 57–65 or aa 52–56 inhibited binding of eNAMPT to TLR4 or CYBB in macrophages. Consistent with the blockade of iNAMPT– TLR4 by TAT–NAMPT (aa 57–65) and iNAMPT–CYBB by TAT–NAMPT (aa 52–56), TAT– NAMPT (aa 57–65) specifically inhibited eNAMPT–TLR4 binding, and TAT–NAMPT (aa 52–56) specifically inhibited eNAMPT–CYBB binding (Figure 3E). Moreover, TAT– NAMPT (aa 52–65) inhibited the eNAMPT–TLR4 and eNAMPT–CYBB binding interactions. These findings show that NAMPT aa 57–65 and aa 52–56 are necessary for interacting with TLR4 or CYBB, respectively, demonstrating that NAMPT interactions with TLR4 and CYBB are genetically separate.
我们接下来测试了TAT-NAMPT AA 57-65或AA 52-56是否抑制巨噬细胞中eNAMPT与TLR4或CyBB的结合。与TAT-NAMPT(AA 57-65)和TAT-NAMPT(AA 52-56)阻断iNAMPT-TLR4和iNAMPT-CyBB一致,TAT-NAMPT(AA 57-65)特异性抑制eNAMPT-TLR4结合,TAT-NAMPT(AA 52-56)特异性抑制eNAMPT-CYBB结合(图3e)。此外,TAT-NAMPT(AA 52-65)抑制eNAMPT-TLR4和eNAMPT-CyBB的结合作用。这些发现表明,NAMPT AA 57-65和AA 52-56分别是与TLR4或CyBB相互作用所必需的,表明NAMPT与TLR4和CyBB的相互作用在基因上是分开的。
3.4. NAMPT多肽抑制NLRP3炎症体的激活
To determine the effect of NAMPT peptides related to signal 1 for the NLRP3 inflammasome in macrophages, we exposed LPS-treated BMDMs to different concentrations of NAMPT peptides (aa 57–65, aa 52–56, or aa 52–65). NAMPT peptides aa 57–65 and aa 52–56 partially inhibited the activation of NF-κB, generation of ROS, and production of cytokines; however, NAMPT peptides aa 52–65 markedly inhibited the generation of ROS and NF-κB-induced cytokine production (Figure 4A–C). Further, TAT–NAMPT peptides aa 52–65, but not TAT–NAMPT peptides aa 57–65 and aa 52–56, partially inhibited NLRP3 inflammasome activation signal 1 (Figure S2A). Thus, the TAT–NAMPT peptide was less likely to inhibit the interaction of eNAMPT with TLR4 or CYBB
为了确定与信号1相关的NAMPT多肽对巨噬细胞中NLRP3炎症小体的影响,我们将脂多糖处理的BMDM暴露于不同浓度的NAMPT多肽(AA 57-65、AA 52-56或AA 52-65)。NAMPT多肽AA 57-65和AA 52-56部分抑制了NF-κB的激活、ROS的产生和细胞因子的产生;但AA52-65显著抑制ROS的产生和NF-κB诱导的细胞因子的产生(图4A-C)。此外,TAT-NAMPT多肽AA 52-65,而不是TAT-NAMPT多肽AA 57-65和AA 52-56,部分抑制了NLRP3炎症体激活信号1(图S2A)。因此,TAT-NAMPT多肽不太可能抑制eNAMPT与TLR4或CyBB的相互作用。
图4:NAMPT多肽抑制NLRP3炎症体的信号1和2的激活。(A-C)BMDM用内毒素刺激4h,用rNAMPT预处理1h(A、A、B)或18h(C)。(A)将WCL与αiKb-α,αP-ikb-α或α肌动蛋白一起用于IB。(B)流式细胞仪分析过氧化氢(20,70-二氯二乙酸酯的探针)或O2-(二氢乙锭的探针)。定量分析H_2O_2和O_2-(上)的平均荧光强度。NADPH氧化酶活性(低)。(C)收集培养上清液,进行细胞因子酶联免疫吸附试验。(d,E)脂多糖刺激的BMDM先用rNAMPT处理1h,再用ATP激活30min。(D)IL-1β和IL-18的酶联免疫吸附试验。(E)使用αIL-1βp17、αIL-18 p18或αCASP1 p10的SN中的IB,以及使用αPro-IL-1、β,αPro-IL-18、αPro-CASP1或α肌动蛋白的WCL。这些数据来自七个独立的实验,它们产生了可比的结果(A,E)。这些数据来自三个独立的实验,它们产生了类似的结果。(B-D)。给出了三个实验的均数±标准差。与内毒素+PBS相比有显著差异(*p<0.001
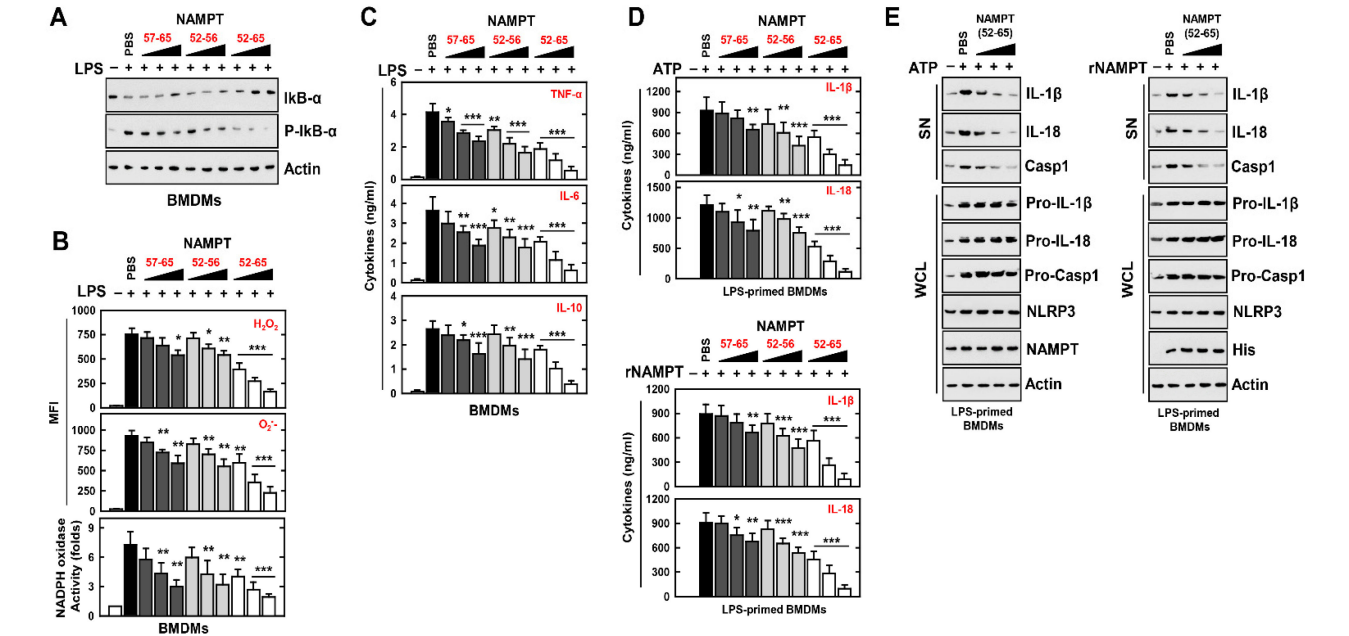

图S2:TAT-NAMPT或NAMPT多肽对炎症反应的影响(A)BMDM用内毒素(100 ng/mL)刺激18h,再用rNAMPT预处理1h。收集培养上清液,进行细胞因子酶联免疫吸附试验。(B)脂多糖刺激的BMDM先用rNAMPT处理1h,然后用尼格列星激活30min或DSS激活24h。酶联免疫吸附试验检测IL-1、β和IL-18。这些数据代表了三个具有类似结果的独立实验。所显示的数据是三个实验的平均值±SD。与内毒素+PBS组比较,差异有统计学意义(P<0.001)。
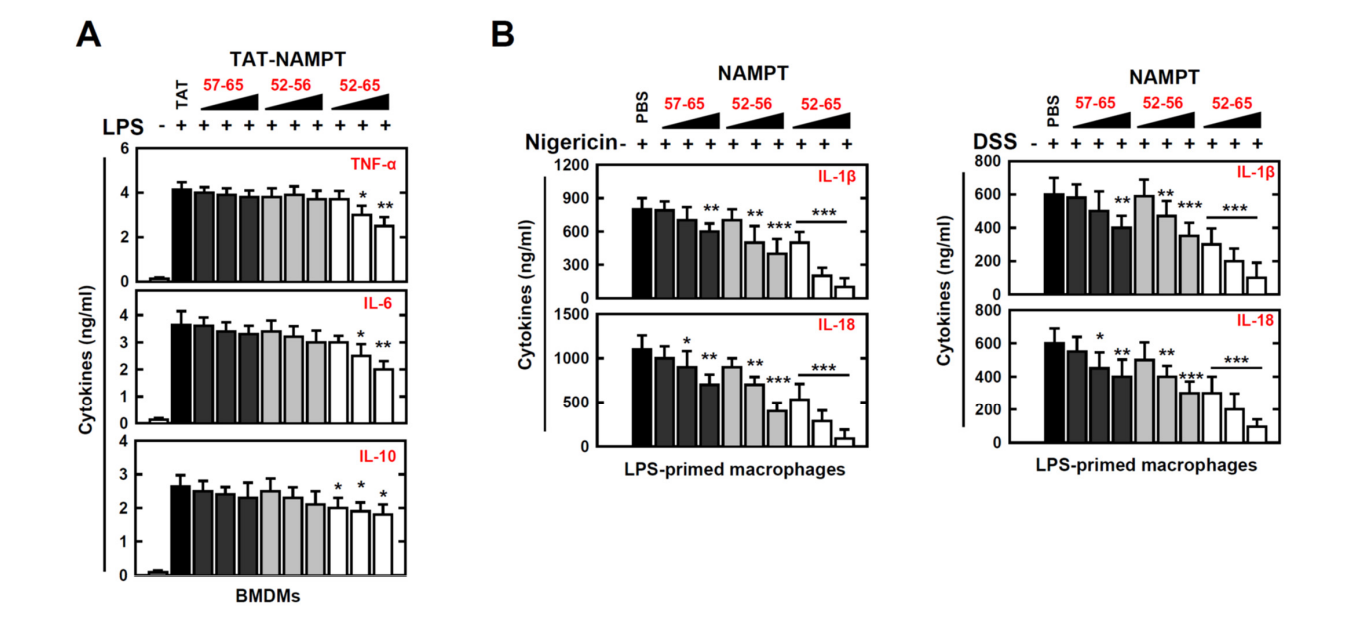

We therefore explored if NAMPT peptides have a special role in the modulation of signal-2 activation of the NLRP3 inflammasome. We found that NAMPT peptide aa 52–65 efficiently inhibited the maturation of IL-1β and IL-18, as well as ATP-induced caspase-1 cleavage and nigericin or DSS stimulation (Figure 4D upper, Figure 4E left and Figure S2B) We found that the NAMPT peptide aa 52–65 specifically inhibited the action of eNAMPT (Figure 4D lower, Figure 4E right). These findings indicate that NAMPT’s actions in the TLR4-mediated signaling pathway and in CYBB-containing NOXs are functionally and genetically separable. Together, these data provide evidence that the NAMPT peptide aa 52–65 is an essential negative regulator of signal 1 and signal 2 in response to NLRP3 inflammasome activation.
因此,我们探索了NAMPT多肽是否在NLRP3炎症小体的信号-2激活的调制中具有特殊的作用。我们发现,NAMPT多肽AA 52-65能有效地抑制IL-1β和IL-18的成熟,以及对caspase-1的裂解和黑素或DSS的刺激(图4D,图4E左侧和图S2B)。这些发现表明,NAMPT在TLR4介导的信号通路中和在含有CyBB的NOx中的作用在功能和基因上是可以分离的。综上所述,这些数据提供了证据,证明NAMPT多肽AA 52-65是NLRP3炎症小体激活反应中信号1和信号2的重要负调节因子。
3.5. 针对病理性炎性结肠炎的重组多功能CT-NAMPT蛋白
The experiments presented above indicate that NAMPT peptides (aa 57–65 and aa 52–56) directly blocked the binding of eNAMPT to the extracellular domains of CYBB and TLR4 and subsequently attenuated the activation of the NLRP3 inflammasome in macrophages. We used the peptide TWYKIAFQRNRK (designated TK), derived from the COOH-terminal globular domain of laminin-1 α1 chain, as a vehicle for targeted drug delivery to the colon. Thus, TK interacts with integrin α6β1, with high affinity for colonic tissue [23,41]. Accordingly, we developed a recombinant 12-residue TK peptide (CT) conjugated to a multifunctional NAMPT (rCT-NAMPT) in which CT targets the colon and harbors the essential and minimal aa residues required for CYBB/TLR4 binding. The authenticity of the predicted product was confirmed using SDS–polyacrylamide gel electrophoresis and immunoblotting (Figure 5A left). There were no significant differences compared with the vehicle control associated with rCT-NAMPT-induced cytotoxicity in BMDMs (Figure 5A right).
上述实验表明,NAMPT多肽(AA57-65和AA52-56)可直接阻断eNAMPT与细胞色素P4和TLR4胞外区的结合,从而减弱巨噬细胞中NLRP3炎症体的激活。我们使用多肽TWYKIAFQRNRK(指定为TK)作为靶向药物输送到结肠的载体,TWYKIAFQRNRK来自层粘连蛋白-1α1链的COOH端球状结构域。因此,TK与整合素α6β1相互作用,对结肠组织具有高亲和力[23,41]。因此,我们开发了一种与多功能NAMPT(RCT-NAMPT)偶联的重组12个残基TK肽(RCT-NAMPT),其中CT以结肠为靶标,并含有CYBB/TLR4结合所需的基本和最低AA残基。用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳法和免疫印迹法证实了预测产物的真实性(图5A左)。与RCT-NAMPT诱导的BMDM细胞毒性相关的赋形剂对照组相比,没有显著差异(图5A右侧)。
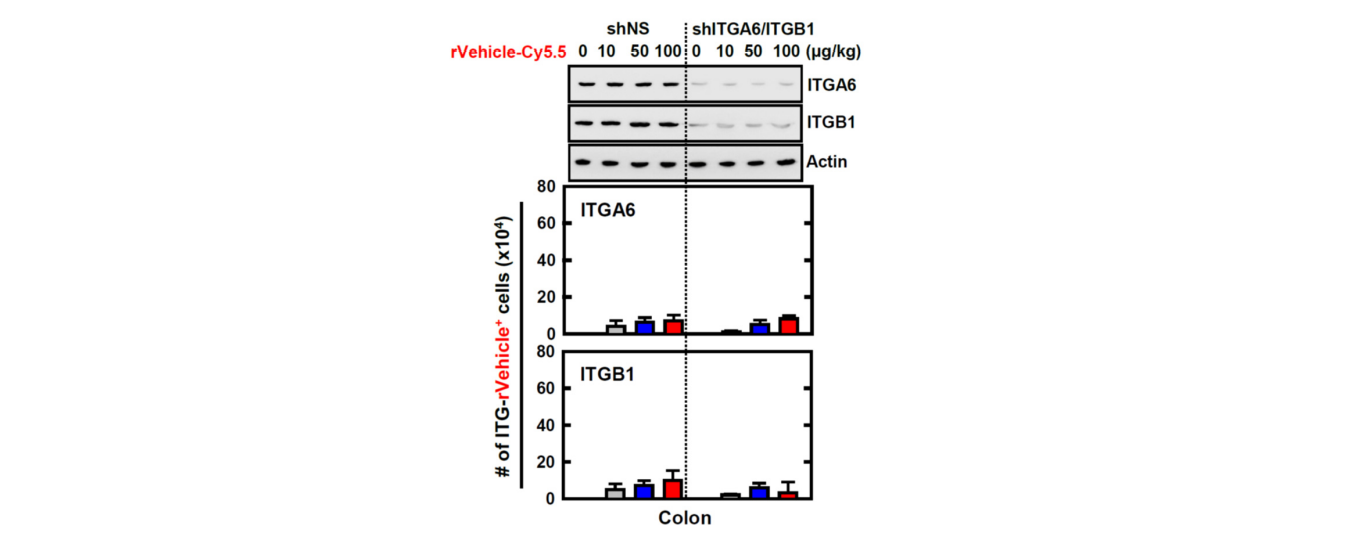
图5:RCT-NAMPT针对发炎的结肠。(A)RCT-NAMPT示意图(上)。经细菌纯化的6xHis-rct-NAMPT、rVehicle或rCT用考马斯蓝染色或IB加αHis(左下)分析。用rVehicle、RCT或RCT-NAMPT处理BMDM,按照所述的时间和浓度进行处理,并用四甲基偶氮唑盐比色法(下,右)测定细胞存活率。(2)DSS诱导的结肠炎模型。小鼠接受3%DSS连续6天,并在第6天(左侧)进行评估。取结肠标本,用α、α、α肌动蛋白进行免疫印迹,用流式细胞仪分析ITGA6+、ITGB1+细胞的数量。(C)Lenti-shNS或Lenti-shITG病毒转导的急性结肠炎模型加3%DSS治疗方案。RCT-NAMPT与Cy5.5联用1h后,取结肠(左)。取结肠标本,分别加入α、α、α肌动蛋白,用流式细胞仪分析ITGA6+或ITGB1+细胞数。这些数据来自三个独立的实验,它们产生了可比较的结果(A-C)
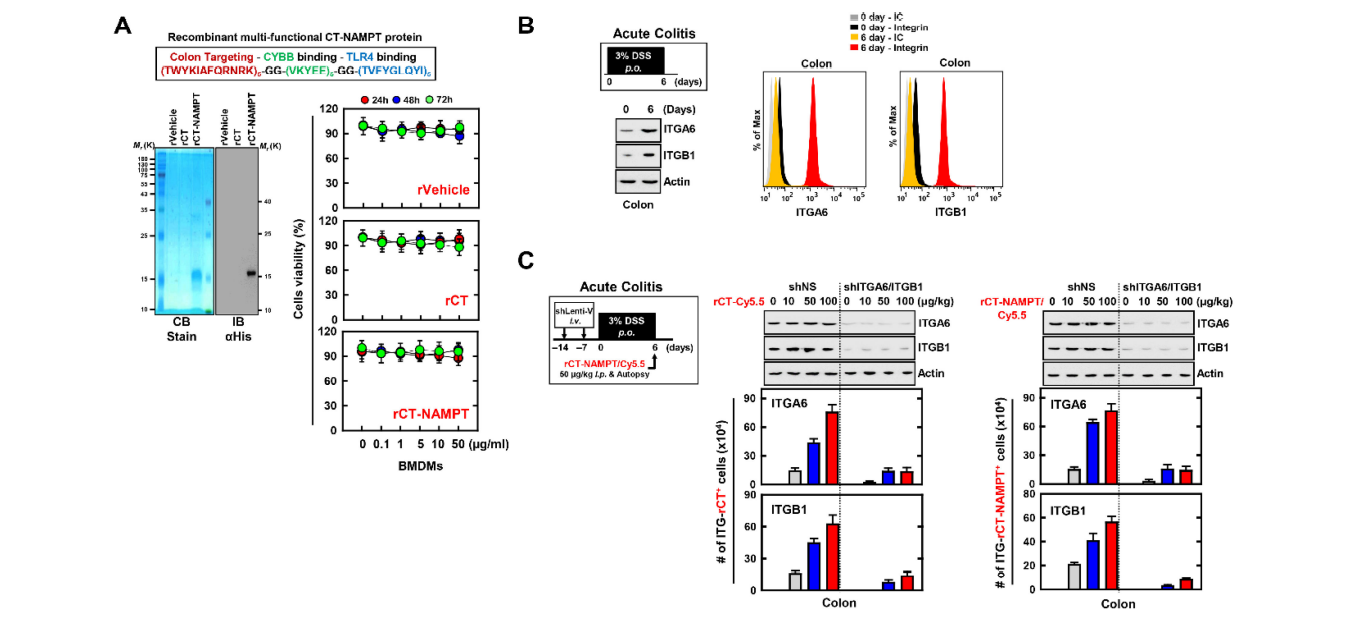

We employed a mouse model of DSS-induced colitis to further investigate the physiological significance of rCT-NAMPT in inflammatory colitis (Figure 5B,C). We found that the expression of ITGA6 (integrin α6) and ITGB1 (integrin β1), which bind to TK peptides (CT), was significantly increased in the colon of mice with acute colitis (Figure 5B). Next, to evaluate the specificity of rCT-NAMPT, we generated ITGA6- or ITGB1-knockdown mice through sh-Lentiviral transduction. The mice were treated with DSS and adminis- tered rCT-NAMPT/Cy5.5 via intraperitoneal injection on day 6. The rCT or rCT-NAMPT specifically targeted colonic tissues in mice with acute colitis, but not in other organs, in a concentration-dependent manner (Figure 5C and Figure S3, and data not shown). These results show that rCT binds to colonic tissues, which raises the possibility of designing colon-targeted drug delivery systems for use as pharmaceutical applications for treating DSS-induced acute colitis.
我们采用DSS诱导的小鼠结肠炎模型来进一步研究RCT-NAMPT在炎症性结肠炎中的生理学意义(图5B,C)。我们发现,在急性结肠炎小鼠的结肠中,与TK肽(CT)结合的整合素α6和整合素β1的表达显著增加(图5B)。接下来,为了评估RCT-NAMPT的特异性,我们通过sh-慢病毒转导获得了ITGA6或ITGB1基因敲除的小鼠。第6天给予DSS和RCT-NAMPT/Cy5.5腹腔注射。RCT或RCT-NAMPT以浓度依赖的方式针对急性结肠炎小鼠的结肠组织,而不是其他器官(图5C和图S3,数据未示出)。这些结果表明,RCT与结肠组织结合,这增加了设计结肠靶向给药系统用于治疗DSS诱导的急性结肠炎的药物应用的可能性。
图S3:RVehicle肽对结肠靶向实验条件的影响遵循图2中概述的方案。用α、α、α肌动蛋白作免疫印迹,用流式细胞仪分析ITGA6+、ITGB1+细胞数。这些数据代表了三个具有类似结果的独立实验。

3.6. RCT-NAMPT对DSS诱导的小鼠急慢性结肠炎的缓解作用
We next evaluated the medicinal effects of rCT-NAMPT on mouse models in DSSinduced acute and chronic colitis. For this purpose, we generated ITG (ITGA6 and ITGB1)knockdown mice through sh-Lentiviral transduction on days -7 and -14 via i.v. before DSS treatment. While the mice were treated with 3% DSS for 6 days, rVehicle, rCT or rCT-NAMPT (50 µg/kg) was i.p. injected 8 times. rCT-NAMPT significantly increased the survival rates of Lenti-shNS-transduced mice with DSS-induced colitis, but not those of Lenti-shITG-transduced mice. Neither rVehicle or rCT detectably affected mortality, suggesting that NAMPT peptides contribute to the regulation of the inflammatory response (Figure 6A). Furthermore, body-weight loss in Lenti-shNS-transduced mice treated with rCT-NAMPT was reduced by roughly 20% when compared to rVehicle- or rCT-treated mice. Body weights of Lenti-shITG-transduced mice were higher than in Lenti-shNS-transduced mice, but there was no significant difference (Figure 6B).
接下来,我们评估了RCT-NAMPT在DSS诱导的急性和慢性结肠炎小鼠模型上的药效。为此,我们通过静脉注射在第-7天和第-14天通过慢病毒转导获得了ITG(ITGA6和ITGB1)基因敲除小鼠。在DSS治疗前。3%DSS灌胃6d后,rVehicle、RCT或RCT-NAMPT(50µg/kg)腹腔注射。注射了8次。RCT-NAMPT可显著提高Lenti-shNS转基因小鼠DSS诱导的结肠炎的存活率,但对Lenti-shITG转基因小鼠的存活率无明显影响。RVehicle或RCT都不能检测到对死亡率的影响,这表明NAMPT多肽有助于调节炎症反应(图6A)。此外,与rVehicle或RCT治疗的小鼠相比,接受RCT-NAMPT治疗的Lenti-shNS转基因小鼠的体重损失减少了约20%。转Lenti-shITG基因小鼠的体重高于转Lenti-shNS基因小鼠,但差异不显著(图6B)。
图6.RCT-NAMPT对DSS诱导的小鼠急性结肠炎有药效。(A)Lenti-shNS或Lenti-shITG病毒转导的急性结肠炎模型,给予3%DSS+RCT-NAMPT(50微克/公斤)(上)。监测小鼠的存活时间为12天;每组n=15只小鼠(下限)测量死亡率。RVehicle处理的动物之间的统计差异被注意到(LOG-RANK检验)。这些数据来自两个不同的实验,它们产生了类似的结果。(B)体重减轻(n=8)。(C)根据临床参数(体重减轻、大便稠度、出血)获得结肠炎评分(n=8)。(D)用rVehicle、RCT或RCT-NAMPT治疗3%DSS组小鼠(n=8)的结肠图像(上)和长度(下)。(E)使用冒号与αHis或αNAMPT一起IP,然后IB与αTLR4、αCybb或αCyba一起使用。用α-TLR4、α-CYBB、α-CYBA、α-His或α-肌动蛋白作为IB。(F)结肠细胞ROS的流式细胞仪分析(n=8)。(G)结肠组织匀浆中细胞因子水平和MPO活性(n=10)。(H)结肠(左侧)苏木精-伊红(H&E)染色(n=10):按照技术(材料和方法)中的指示,使用rVehicle、RCT或RCT-NAMPT对3%DSS处理的小鼠进行典型的影像组织病理学评分。与rVehicle相比,使用带Bonferroni校正的学生t检验(*p<0.001)来评估统计学意义。
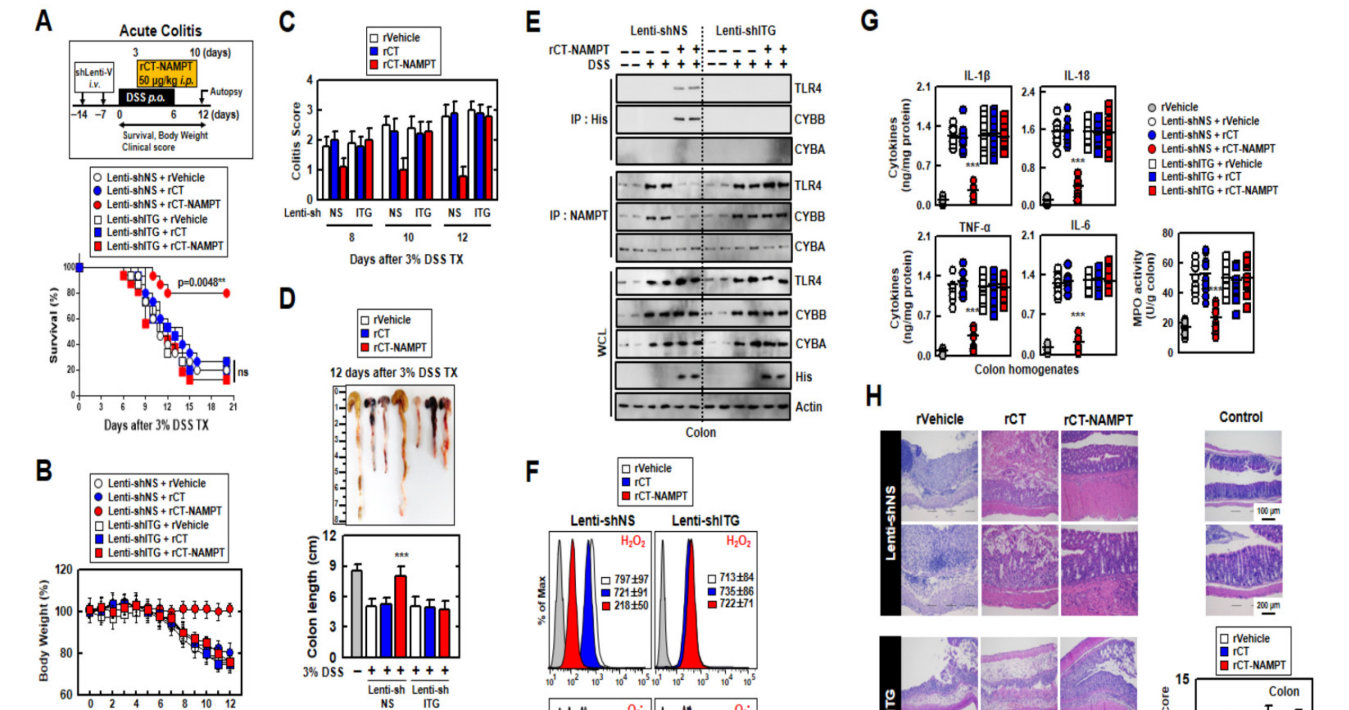
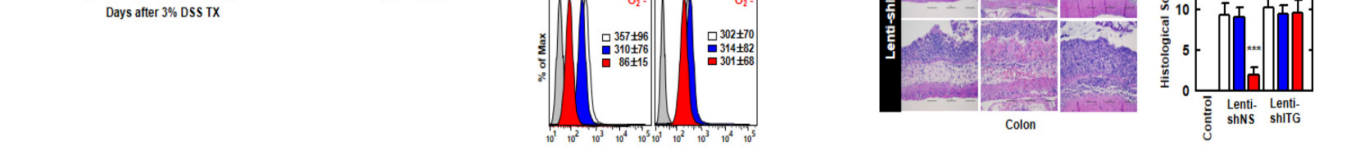

The colitis scores of mice were markedly decreased in Lenti-shNS-transduced mice treated with rCT-NAMPT (Figure 6C). After 12 days, we measured the length of the colon, which is an indication of colitis. Colon length recovered in rCT-NAMPT-treated Lenti-shNS-transduced mice, but remained unchanged in Lenti-shITG-transduced mice (Figure 6D). Further, we tested whether rCT-NAMPT exerted pharmacological activity in vivo. For example, the in vivo detection of interactions between the NAMPT and TLR4mediated signaling pathways and CYBB-containing NOXs may prove to be important for the evaluation of rCT-NAMPT to identify drugs that treat lethal inflammatory disease. For this purpose, we analyzed the binding of NAMPT, ROS levels, and cytokine production in the colon. The interaction of NAMPT with TLR4 or CYBB was detected only in the colon of DSS-treated Lenti-shNS-transduced mice, but not in Lenti-shITG-transduced mice (Figure 6E). Cellular ROS levels decreased in Lenti-shNS-transduced mice treated with rCT-NAMPT, but not in Lenti-shITG-transduced mice (Figure 6F).
经RCT-NAMPT治疗的Lenti-shNS转基因小鼠的结肠炎评分显著降低(图6C)。12天后,我们测量了结肠的长度,这是结肠炎的一个迹象。RCT-NAMPT治疗的Lenti-shNS转导小鼠的结肠长度恢复,但Lenti-shITG转导小鼠的结肠长度保持不变(图6D)。此外,我们还测试了RCT-NAMPT在体内是否具有药理活性。例如,体内检测NAMPT和TLR4介导的信号通路与含有CYBB的NOx之间的相互作用可能被证明对于评估RCT-NAMPT以识别治疗致死性炎症疾病的药物是重要的。为此,我们分析了NAMPT、ROS水平和细胞因子在结肠中的结合。NAMPT与TLR4或CyBB的相互作用仅在DSS处理的Lenti-shNS转导小鼠的结肠中检测到,而在Lenti-shITG转导的小鼠中未检测到(图6e)。使用RCT-NAMPT治疗的Lenti-shNS转导小鼠的细胞ROS水平降低,但Lenti-shITG转导小鼠的细胞ROS水平没有下降(图6F)。
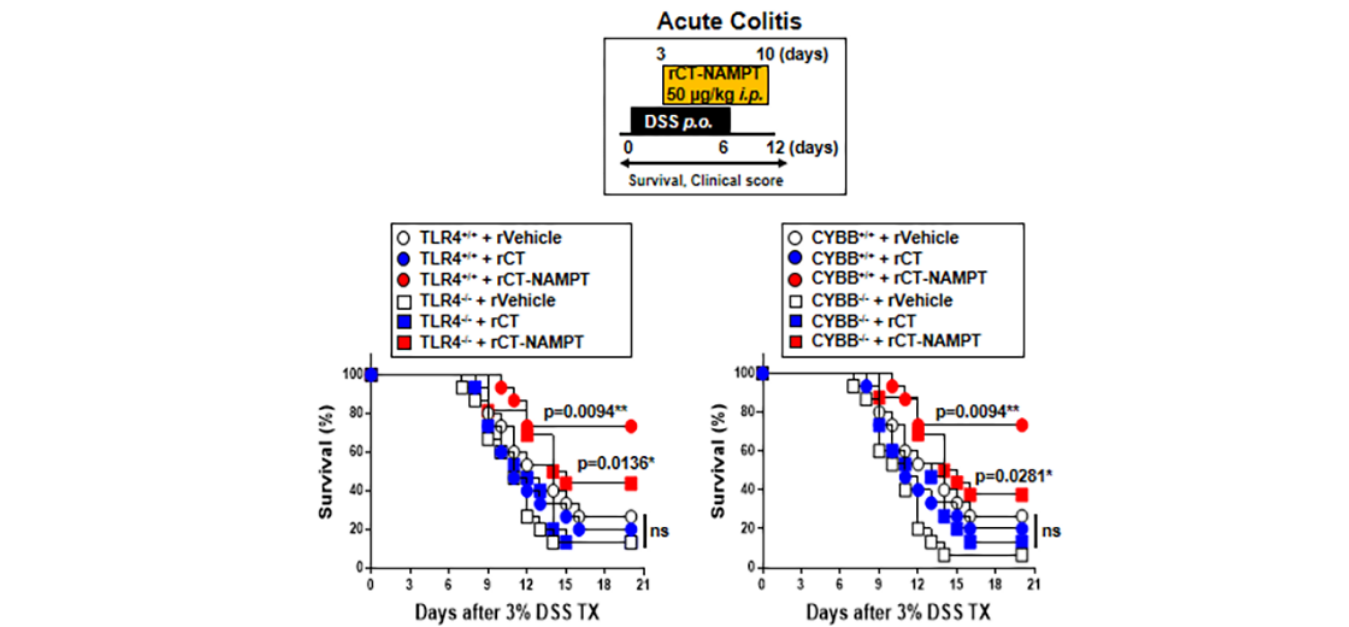
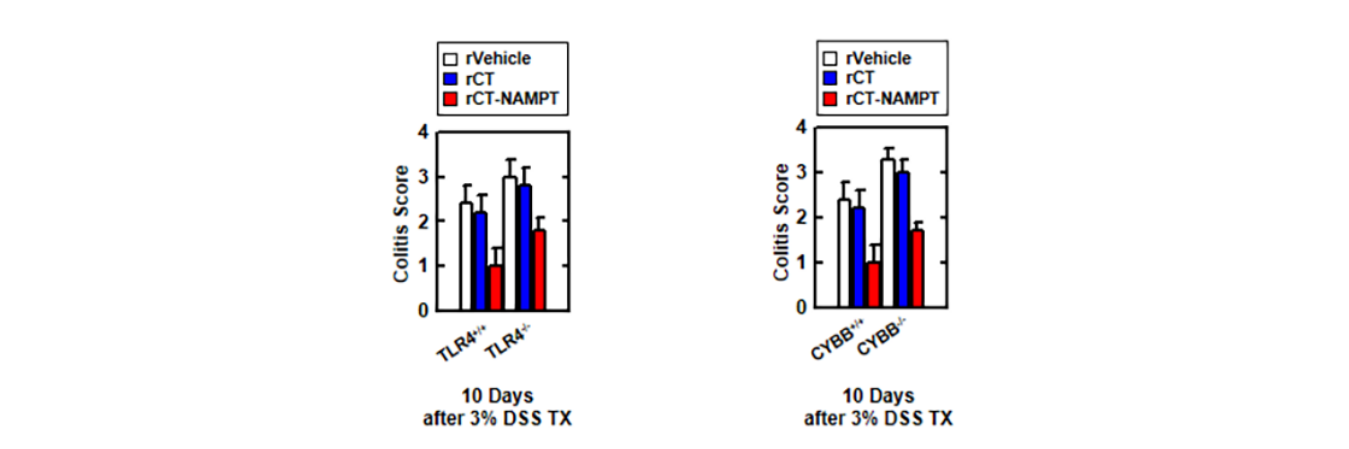
Moreover, the activity of myeloperoxidase involved in the production of TNF-α, IL-1β, IL-6, and IL-18 as well as the activation of NLRP3 inflammasome was measured (Figure 6G). The histological scores of colitis (H&E staining) revealed that the rVehicle- and the rCTtreated colon was disrupted by DSS in Lenti-shNS-transduced mice, and the addition of rCT-NAMPT restored the colonic barrier. Further, ITGA6/ITGB1-knockdown had no effect on rCT-NAMPT because of loss of colon-targeting ability (Figure 6H). Notably, rCT-NAMPT has a partially therapeutic effect against acute DSS-induced colitis in TLR4−/− or CYBB−/− mice compared with WT mice (Figure S4).
此外,还检测了参与肿瘤坏死因子-α、IL-1β、IL-6和IL-18的产生以及NLRP3炎症小体激活的髓过氧化物酶的活性(图6G)。结肠炎的组织学评分(H&E染色)显示,DSS破坏了Lenti-shNS转基因小鼠的rCT和rCT治疗后的结肠,RCT-NAMPT的加入恢复了结肠屏障。此外,ITGA6/ITGB1-基因敲除对RCT-NAMPT没有影响,因为失去了结肠靶向能力(图6H)。值得注意的是,与WT小鼠相比,RCT-NAMPT对TLR4−/−或CyBB−/−小鼠的急性DSS诱导的结肠炎有部分治疗作用(图S4)。
图S4:RCT-NAMPT对DSS诱导的TLR4-/-和CyBB-/-小鼠急性结肠炎有治疗作用。RCT-NAMPT(50μg/kg)治疗3%DSS慢性结肠炎模型示意图(上)。监测小鼠的存活12天;每组15只小鼠的死亡率。与rVehicle处理的小鼠相比,有统计学差异(对数等级检验)。这些数据代表了两个结果相似的独立实验。根据临床参数(体重减轻、大便稠度、出血)获得结肠炎评分(n=8,较低)。

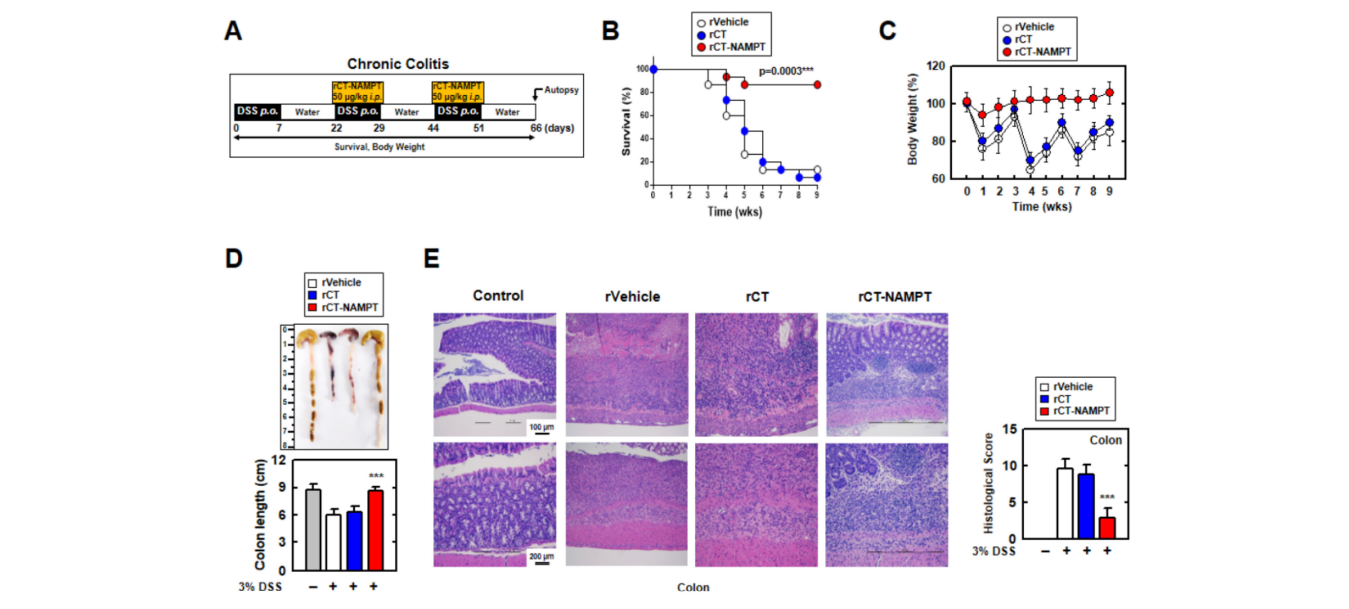
We then investigated the therapeutic effect of rCT-NAMPT on chronic colitis, subjecting mice to DSS and rCT-NAMPT for 66 days (Figure 7A). rCT-NAMPT, like acute colitis, boosted mouse survival rates by roughly 90%. (Figure 7B). Body weights fluctuated in the rVehicle- and rCT-treated mice, although rCT-NAMPT-treated mice maintained their body weights (Figure 7C). Further, the lengths of the colons of rCT-NAMPT-treated mice were restored compared with those of DSS-treated mice, which were significantly damaged (Figure 7D). H&E colon examination demonstrated that rCT-NAMPT dramatically improved colon vitality in DSS-treated mice (Figure 7E). These findings suggest that rCTNAMPT has a therapeutic effect against DSS-induced colitis through inhibiting the NAMPT– TLR4 or –CYBB interaction in vivo.
然后,我们研究了RCT-NAMPT对慢性结肠炎的治疗效果,给小鼠服用DSS和RCT-NAMPT 66天(图7A)。与急性结肠炎一样,RCT-NAMPT将小鼠的存活率提高了约90%。(图7B)。在rVehicle和RCT治疗的小鼠中,体重波动,尽管RCT-NAMPT治疗的小鼠保持其体重(图7C)。此外,与显著受损的DSS组相比,RCT-NAMPT组小鼠的结肠长度得到了恢复(图7D)。H&E结肠检查显示,RCT-NAMPT显著提高了DSS治疗小鼠的结肠活力(图7E)。这些结果提示,rCTNAMPT在体内通过抑制NAMPT-TLR4或-CyBB的相互作用而对DSS诱导的结肠炎有治疗作用。
图7.RCT-NAMPT缓解DSS诱导的小鼠慢性结肠炎。(A)慢性结肠炎模型,给予3%DSS+RCT-NAMPT(50微克/公斤)。(B)追踪小鼠存活9周,计算每组15只小鼠的死亡率。RVehicle处理的动物之间的统计差异被注意到(LOG-RANK检验)。这些数据来自两个不同的实验,它们产生了类似的结果。(C)rVehicle、RCT或RCT-NAMPT小鼠体重减轻(n=15)。(D)用rVehicle、RCT或RCT-NAMPT观察3%DSS诱导的慢性结肠炎小鼠结肠的图像(上)和长度(下)。(E)结肠(左)H&E染色(n=8):具有代表性的图像。用H&E染色评价rVehicle、RCT、RCT-NAMPT对3%DSS诱导的慢性结肠炎小鼠的组织病理学评分。与rVehicle相比,采用带Bonferroni调整的学生t检验来建立统计学意义(*p<0.001)。

4. 讨论
Here we show that rCT-NAMPT served as an effective novel candidate therapeutic for colitis by modulating the NLRP3 inflammasome-mediated immune signaling system through negative regulation of signals 1 and 2 in response to NLRP3-inflammasome activation. This finding represents a major paradigm shift in the treatment of colitis and an urgently needed therapeutic intervention. The main results of this study are as follows: (1) Analyses of two single-cell RNA-seq datasets, patients with colitis, and mouse models of acute and chronic colitis show that NAMPT significantly contributes to the pathology of UC. (2) Specifically, in macrophages, eNAMPT directly interacted with CYBB causing increased production of ROS and activation of the NLRP3 inflammasome in conjunction with the activation of the TNFα and NF-κB signaling pathways through direct stimulation by the eNAMPT–TLR4 interaction. (3) Amino acid residues aa 57–65 and aa 52–56 in NAMPT were essential for the interaction of NAMPT with TLR4 and CYBB, respectively.(4) A colon-targeting peptide designated TK was designed, and when conjugated to a multifunctional NAMPT peptide, targeted the inflamed colon in vivo. Further, the rCTNAMPT-induced peptide inhibited NLRP3 inflammasome activation in vitro and in vivo.(5) rCT-NAMPT showed potential for treating acute and chronic DSS-induced colitis in mice.Collectively, these observations serve as a proof-of-concept for designing host-oriented therapeutic strategies and further demonstrate an engineered eNAMPT-induced NLRP3 inflammasome network associated with colitis.
在这里,我们表明RCT-NAMPT是一种有效的新的治疗结肠炎的候选药物,它通过负调控NLRP3-炎症体激活的信号1和2来调节NLRP3炎症小体介导的免疫信号系统。这一发现代表了结肠炎治疗的重大范式转变和迫切需要的治疗干预。本研究的主要结果如下:(1)对两个单细胞RNA-SEQ数据集、结肠炎患者和急、慢性结肠炎小鼠模型的分析表明,NAMPT在UC的病理过程中起重要作用。(2)在巨噬细胞中,eNAMPT直接与CyBB相互作用,通过eNAMPT-TLR4相互作用的直接刺激,导致ROS的产生增加和NLRP3炎症体的激活,同时激活肿瘤坏死因子α和核因子-κB信号通路。(3)NAMPT中的氨基酸残基AA 57-65和AA 52-56分别是NAMPT与TLR4和CyBB相互作用所必需的。(4)设计了一种结肠靶向多肽TK,并与多功能NAMPT多肽结合,在体内靶向炎症的结肠。此外,rCTNAMPT诱导的多肽在体外和体内都抑制了NLRP3炎症体的激活。(5)RCT-NAMPT显示出治疗DSS诱导的小鼠急性和慢性结肠炎的潜力。
The present data contribute several key insights into the role and therapeutic perspectives of macrophage-derived eNAMPT within the inflammatory response, particularly in the context of IBD. First, we found that macrophages distributed throughout the body, in contrast to T cells, serve as gatekeepers of tissue homeostasis and mediate innate and adaptive immune responses [42,43]. The present analyses of single-cell RNA-seq datasets show that compared to activated macrophages, T cells may be deactivated as suggested by (1) the overpopulation of regulatory T cells and (2) negative enrichment scores for most of the inflammatory signaling pathways in T cells. It is unclear if deactivation of T cells led to overactivated macrophages, or the opposite. Further, we are unable to explain why P2RY6 was not detectably or infrequently expressed in macrophages derived from inflamed UC samples. Nevertheless, blockade of the eNAMPT–CYBB interaction, reduction of ROS production, or both, in macrophages may ameliorate UC by relieving the activation of NLRP3 inflammasome. These two issues require further in-depth studies to enhance our knowledge of the pathogenesis of UC and other autoimmune diseases.
目前的数据为巨噬细胞来源的eNAMPT在炎症反应中的作用和治疗前景提供了几个关键的见解,特别是在IBD的背景下。首先,我们发现,与T细胞不同,巨噬细胞分布于全身,是组织内稳态的守门人,并介导先天和获得性免疫反应[42,43]。目前对单细胞RNA-seq数据集的分析表明,与激活的巨噬细胞相比,T细胞可能失活,原因如下:(1)调节性T细胞数量过多,(2)T细胞中大多数炎症信号通路的浓缩分数为负。目前尚不清楚T细胞的失活是导致巨噬细胞过度激活,还是相反。此外,我们无法解释为什么P2RY6在发炎的UC样本中的巨噬细胞中没有检测到或很少表达。然而,阻断eNAMPT-CYBB相互作用,减少巨噬细胞内ROS的产生,或两者兼而有之,可能通过减轻NLRP3炎症体的激活而改善UC。这两个问题需要进一步深入研究,以提高我们对UC和其他自身免疫性疾病的发病机制的认识。
Second, this is the first report, to our knowledge, showing that macrophage-specific eNAMPT functions as a protective factor to alleviate the severity of colitis through inhibiting the binding by macrophages of CYBB and TLR4 to circulating eNAMPT. Recent studies have shown that NAMPT produced by macrophages binds to the C-C motif chemokine receptor type 5 in vivo, providing a stem cell activation niche that promotes muscle repair and regeneration [44]. Further, inhibition of NAMPT by the pharmacological inhibitor FK866 or genetic ablation of Nampt (Nampt mKO) may abrogate proliferation-inducing cues required for the injury-induced repair response in DSS-induced colitis [8]. However, as mentioned in this study [8], whether eNAMPT secreted by macrophages promotes colon repair and regeneration has not been determined. These findings [8], together with the comprehensive evidence of the present study, support the conclusion that macrophagespecific eNAMPT alleviates the severity of colitis. The preclinical work of Colombo et al underlines that eNAMPT has a role in the etiology of some inflammatory illnesses via the effect of eNAMPT on macrophage inflammation. Furthermore, RNA-seq investigation of peritoneal macrophages reveals that eNAMPT activates an M1-distorted transcriptional program and preferentially promotes IFN-driven transcriptional activation in macrophages and human monocyte-derived macrophages via STAT1/3 phosphorylation [43].
第二,据我们所知,这是第一份报告,表明巨噬细胞特异性eNAMPT通过抑制巨噬细胞与循环eNAMPT的结合而发挥保护因子的作用,以减轻结肠炎的严重程度。最近的研究表明,巨噬细胞产生的NAMPT在体内与C-C基序趋化因子受体5结合,提供干细胞激活的利基,促进肌肉修复和再生[44]。此外,用药物抑制剂FK866抑制NAMPT或基因消融NAMPT(NAMPT MKO)可能会消除DSS诱导的结肠炎损伤诱导修复反应所需的增殖诱导信号[8]。然而,正如本研究[8]所提到的,巨噬细胞分泌的eNAMPT是否促进结肠修复和再生尚未确定。这些发现[8],加上本研究的综合证据,支持巨噬细胞特异性eNAMPT减轻结肠炎严重程度的结论。科伦坡等人的临床前工作强调了eNAMPT通过对巨噬细胞炎症的影响在一些炎症性疾病的病因学中发挥作用。此外,对腹膜巨噬细胞的RNA-SEQ研究表明,eNAMPT激活了M1扭曲的转录程序,并通过STAT1/3磷酸化优先促进巨噬细胞和人单核细胞来源的巨噬细胞中干扰素驱动的转录激活[43]。
Third, we show here that the eNAMPT–CYBB and –TLR4 axes contribute to the pathogenesis of colitis. Increased production of NOXs generates ROS, which are required for phagocytosis [8,32]. In this regard, our present findings highlight the important role of a metabolic axis composed of eNAMPT, NOX and ROS in maintaining the phagocytic activity of macrophages. Further, TLR4 serves as an alternative receptor for eNAMPT, as indicated by SPR studies and then confirmed by additional evidence [17]. For example, in human monocytes, a TLR4-neutralizing antibody reduces eNAMPT-mediated NF-kB activation [18].
第三,我们在这里表明eNAMPT-CYBB和-TLR4轴在结肠炎的发病机制中起作用。NOx的产量增加会产生ROS,这是吞噬细胞所必需的[8,32]。在这方面,我们目前的发现强调了由eNAMPT、NOX和ROS组成的代谢轴在维持巨噬细胞吞噬活性方面的重要作用。此外,TLR4作为eNAMPT的替代受体,如SPR研究所表明的,然后被更多的证据证实[17]。例如,在人类单核细胞中,TLR4中和抗体降低了eNAMPT介导的NF-kB激活[18]。
Molecular docking studies on a minimized model of the NAMPT–TLR4 complex reveal a role for NAMPT-positive patches and the presence of stabilizing electrostatic interactions between K48, K68, K71, K73 with the negatively charged carboxyl moieties of TLR4 [40]. We show here that the amino acid residues 57–65 of NAMPT interacted with the LRR of TLR4. However, recent results demonstrate that eNAMPT exerts IFNγ-induced macrophage polarization independently of the TLR4-independent pathway [43]. KEGG analysis, a previous study, and our present data reveal the consistent enrichment of TLR signaling [45,46]. We therefore believe that it is reasonable to conclude that eNAMPT activity is TLR receptor- and ligand-specific and that the receptor may belong to the TLR family. Therefore, targeting these interactions may represent a promising therapeutic strategy designed to inhibit eNAMPT-induced inflammatory responses.
对NAMPT-TLR4络合物最小模型的分子对接研究揭示了NAMPT阳性斑块的作用以及K48、K68、K71、K73与TLR4[40]的带负电荷的羧基之间存在稳定的静电相互作用。我们在这里证明了NAMPT的57-65个氨基酸残基与TLR4的LRR相互作用。然而,最近的结果表明,eNAMPT施加干扰素γ诱导的巨噬细胞极化不依赖于TLR4途径[43]。KEGG分析,先前的一项研究,和我们目前的数据显示,TLR信号的持续丰富[45,46]。因此,我们认为eNAMPT活性是TLR受体和配体特异性的结论是合理的,该受体可能属于TLR家族。因此,以这些相互作用为靶点可能是一种有希望的治疗策略,旨在抑制eNAMPT诱导的炎症反应。
Fourth, rCT-NAMPT shows promise as a potential therapeutic. For example, numerous studies show that peptide-based biological agents have significant potential as immunotherapeutic agents against inflammatory diseases [30,31,33–35]. Colonic-targeted drug delivery systems for macromolecules may provide therapeutic benefits such as improved patient compliance (due to its painlessness and self-administration) and lower costs. pH-dependent systems, enzyme-inducing systems, receptor-mediated systems, and magnetically driven systems are among the strategies used to achieve more efficient colonic drug delivery for local or systemic effects [24,47].
第四,RCT-NAMPT显示出潜在的治疗前景。例如,大量研究表明,基于多肽的生物制剂作为抗炎症性疾病的免疫治疗剂具有巨大的潜力[30,31,33-35]。结肠靶向大分子给药系统可能提供治疗益处,例如改善患者的依从性(由于其无痛和自我给药)和更低的成本。PH依赖系统、酶诱导系统、受体介导系统和磁驱动系统是用于实现局部或全身效应的更有效的结肠药物输送的策略[24,47]。
Ligands are an essential component of targeted drug delivery systems, and the selection of ligands with high affinity is of considerable interest [23,24,47–49]. Here, we used TK peptides as suitable targeting ligands with potential applications for colon-targeted therapy [23]. Further studies must focus on different approaches to designing colonictargeted drug delivery systems as well as on pharmaceutical applications and formulation technologies.
配体是靶向给药系统的重要组成部分,具有高亲和力的配体的选择是人们非常感兴趣的[23,24,47-49]。在这里,我们使用TK肽作为合适的靶向配体,潜在地应用于结肠靶向治疗[23]。进一步的研究必须侧重于设计结肠靶向给药系统的不同方法,以及药物应用和配方技术。
Notably, these rCT-NAMPTs do not fulfill the requirements of direct NAMPT enzymatic activities, which represent feasible alternatives to the conventional chemotherapy of colitis. Furthermore, the unknown specificities and selectivity of rCT-NAMPTs make it impossible to link their effects to host immune systems, and a shortcoming of an animal experimental model is whether it truly replicates human pathophysiology Other limitations include safety data, potential feasibility, unknown off-target effects, and pharmacokinetics for in vivo proof-of-concept studies.
值得注意的是,这些RCT-NAMPT不满足直接NAMPT酶活性的要求,这是结肠炎常规化疗的可行替代方案。此外,RCT-NAMPTs的特异性和选择性尚不清楚,无法将其作用与宿主免疫系统联系起来,动物实验模型的一个缺点是它是否真的复制了人类的病理生理学。其他限制包括安全性数据、潜在的可行性、未知的非靶点效应和体内概念验证研究的药代动力学。
Our results show that the NAMPT, TLR4, and CYBB genes associated with mouse colitis share functions with those of humans. It is believed that molecular biology and immunological results from mouse model systems can be translated into insights into human biology and health through interpretation based on mouse experimental data. Therefore, additional studies are warranted to establish whether rCT-NAMPTs can be translated to the clinic and whether their effects can be confirmed in individuals with colitis.
我们的结果表明,与小鼠结肠炎相关的NAMPT、TLR4和CYBB基因与人类的功能相同。人们相信,通过基于小鼠实验数据的解释,来自小鼠模型系统的分子生物学和免疫学结果可以转化为对人类生物学和健康的洞察。因此,有必要进行更多的研究,以确定RCT-NAMPTs是否可以移植到临床,以及它们的效果是否可以在结肠炎患者中得到证实。
In conclusion, to effectively maximize the anti-inflammatory effectiveness of immunotherapies targeted to the activated NLRP3 inflammasome, it is important to combine the specificity of the eNAMPT effect on macrophages as well as its efficacy for colonic targeting. The prospects for the treatment of colitis with next-generation peptide immunotherapeutic depend on the results of future translational and clinical trials.
总之,为了有效地最大化针对激活的NLRP3炎症体的免疫治疗的抗炎效果,将eNAMPT对巨噬细胞的特异性与其在结肠靶向的有效性结合起来是很重要的。下一代多肽免疫疗法治疗结肠炎的前景取决于未来翻译和临床试验的结果。

 浙公网安备 33010602011771号
浙公网安备 33010602011771号