【生化代谢基础笔记】RNA 合成
第一节 原核生物转录的模板和酶
一、原核生物转录模板
- 模板链(Template strand) VS 编码链(Coding strand)
-
模板链为合成模板
-
另一股单链为编码链,mRNA 碱基序列与编码链一致
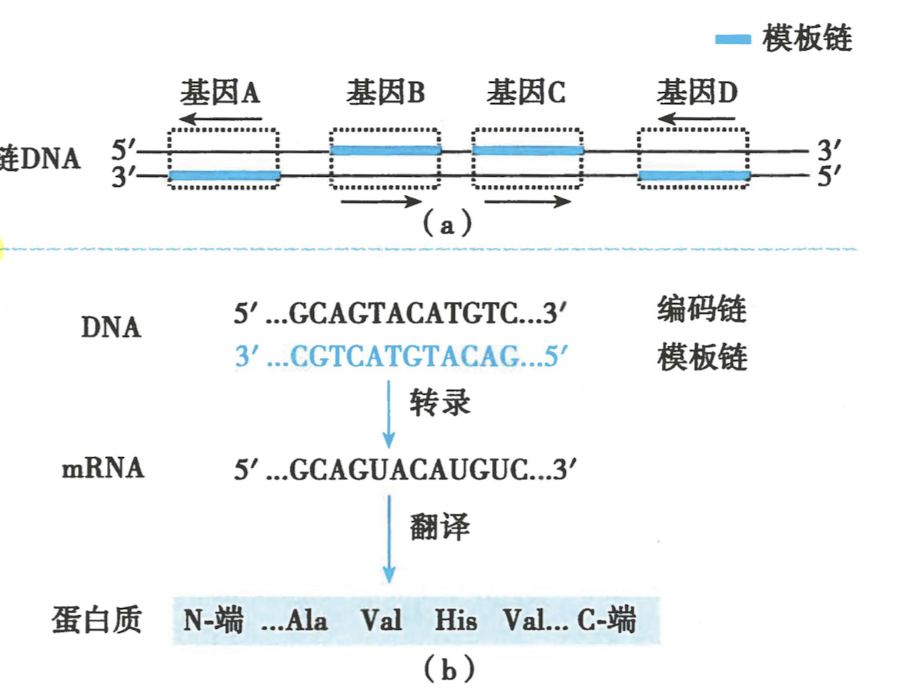
-
二、RNA 聚合酶催化 RNA 的合成
-
RNA 聚合酶能从头启动 RNA 链的合成
-
RNA 聚合酶由多个亚基组成

- 全酶
- σ subunit:辨认转录起始点
- 核心酶(Core enzyme):α2ββ‘ω:转录活性
- HSP 蛋白(热激蛋白)
- 特殊转录起点,σ32 辨认
- 利福平(rifampicin):特异性抑制原核生物 RNA pol的β subunit
- 全酶
三、RNA 聚合酶结合到启动子上启动转录
- 操纵子 Operon:
- 每一段转录区为转录单位
- 包含若干个基因编码区及其调控序列
- 转录起点(TSS/initiator):+1
- 启动子 Promoter(AT rich):
-
-35 区域:RNA pol 对转录起始时的识别序列
-
-10 区域:Pribnow box,形成稳定的 RNA pol-DNA 复合物,转录开始
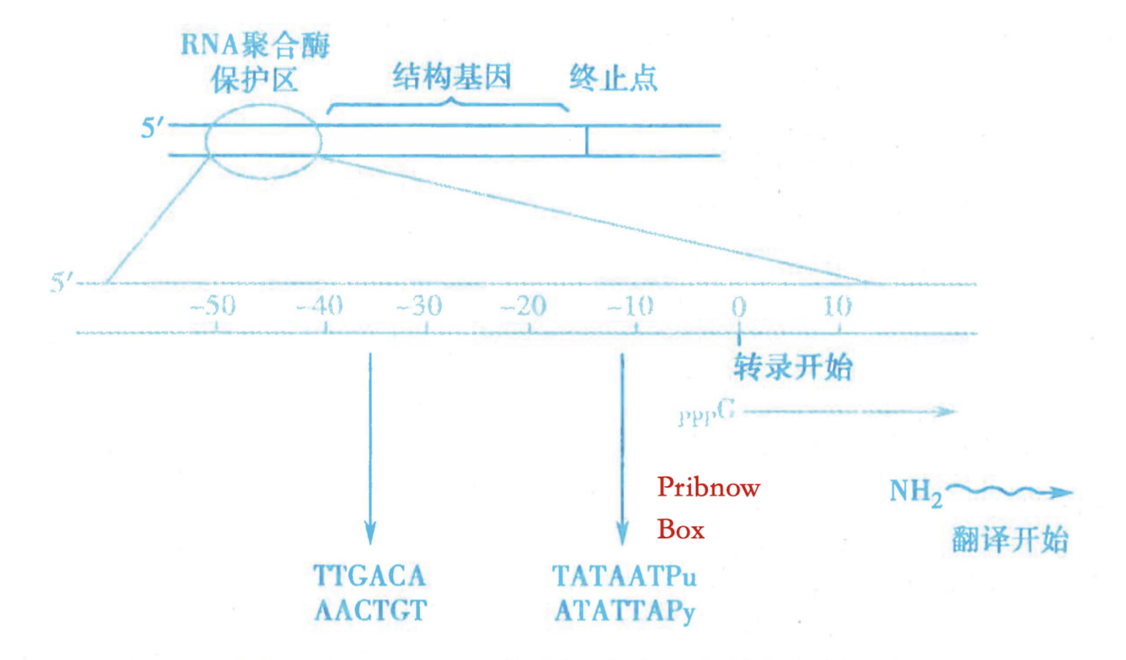
-
第二节 原核生物的转录过程
一、转录起始需要 RNA 聚合酶全酶
- Step1:RNA pol 识别并结合启动子,形成闭合转录复合体
- -35 识别
- 酶移动向-10
- Step2:DNA 双链打开,闭合转录复合体转换为开放转录复合体
- 双链解开范围在 17bp 左右,远小于复制叉
- Step3:第一个磷酸二酯键形成
- 启动子解脱/启动子清除:RNA pol 离开启动子
- 流产式起始(abortive initiation):合成长度小于 10 个核苷酸的 RNA 分子。
二、RNA聚合酶独立延长 RNA 链
-
转录泡
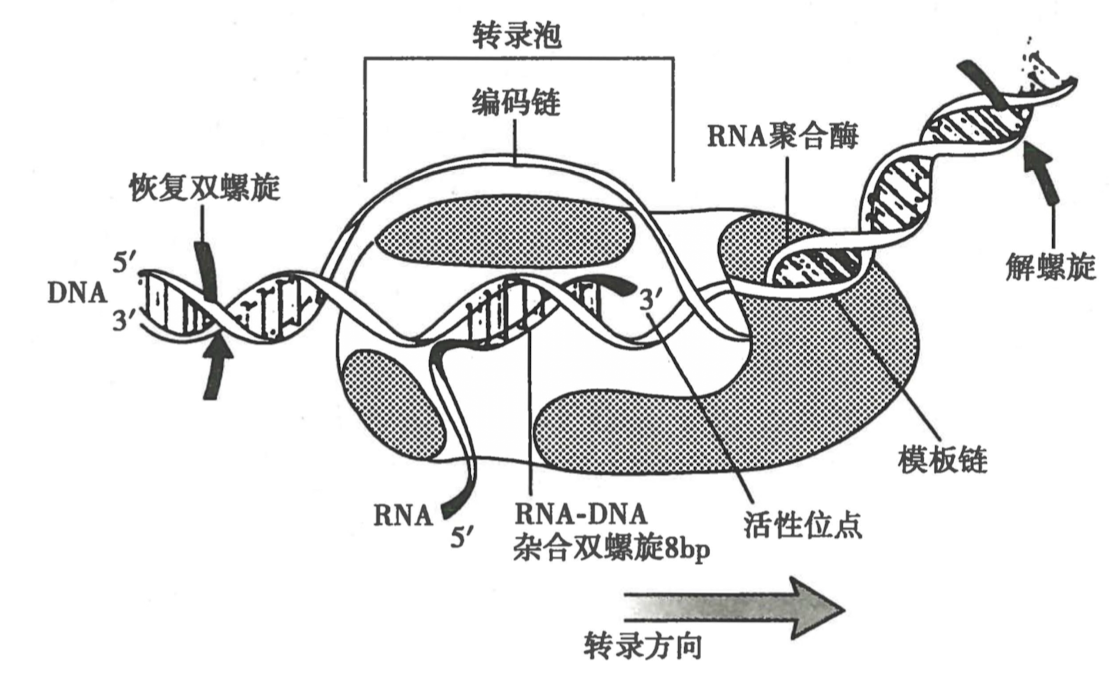
-
转录泡特点:
- 8bp 双螺旋
- 模板链 3-5 = 编码链 5-3
三、原核生物转录延长与蛋白质翻译同时进行
四、原核生物转录终止:ρ-dependent VS ρ-independent
- 依赖 ρ 因子的 termination
-
ρ 因子:
-
同六聚体 pro,亚基 64kD
-
能与 RNA 结合,对 polyC 结合力强
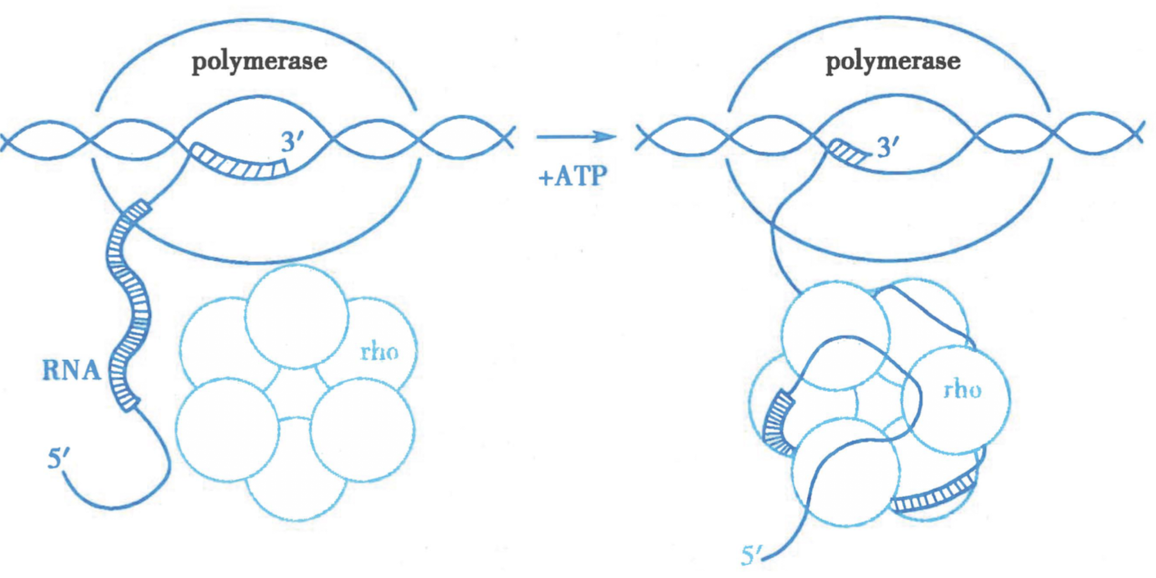
-
-
原理:
- RNA 3‘的 polyC 结合 ρ 因子,RNA pol 移动停止
- ρ有解旋酶活性,解旋 RNA/DNA杂化双链
-
- 非依赖 ρ 因子的 termination
- 3’ polyU 结构,上游GC rich 形成 hairpin 结构
- 机制
-
RNA 分子形成的茎环结构改变 RNA pol 构象
-
poly AU 的不稳定性配对
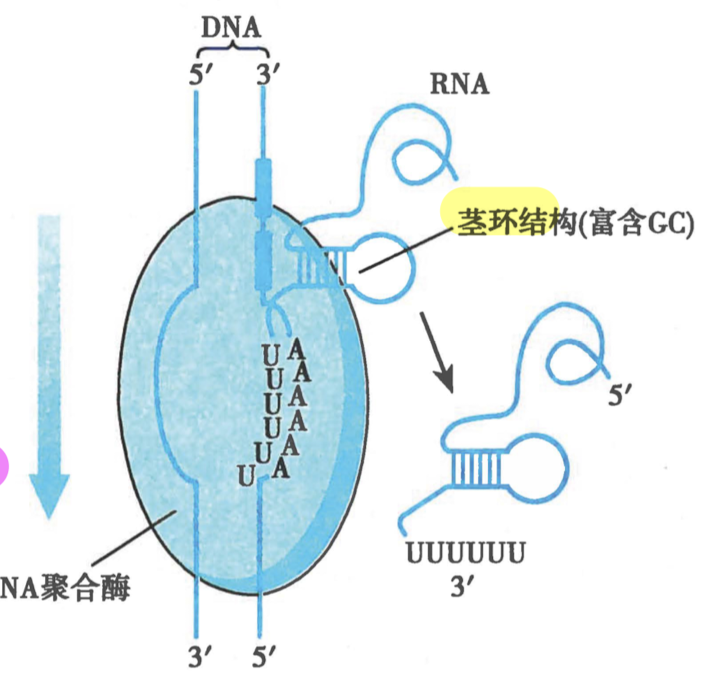
-
第三节 真核生物 RNA 合成
一、真核生物有多重 DNA 依赖的 RNA 聚合酶
| 分类 | 定位 | 功能 | α-鹅膏蕈碱敏感性 |
|---|---|---|---|
| RNA Pol I | 核仁 | 合成 rRNA 前体 | 不敏感 |
| RNA Pol II | 核内 | 合成 mRNA 前体 | 十分敏感 |
| RNA Pol III | 核内 | 合成 tRNA,5SrRNA,核小 RNA | 比较敏感 |
-
结构比原核复杂
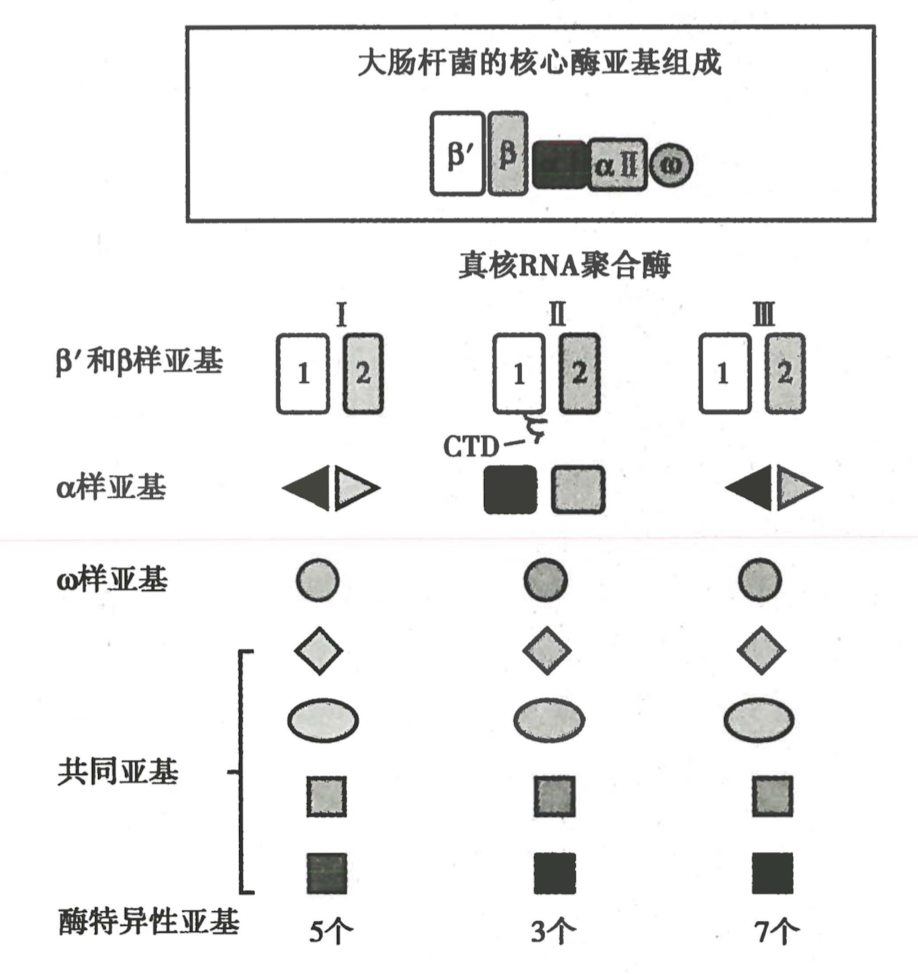
-
Pol II 的 CTD 的磷酸化在转录中起到重要作用
-
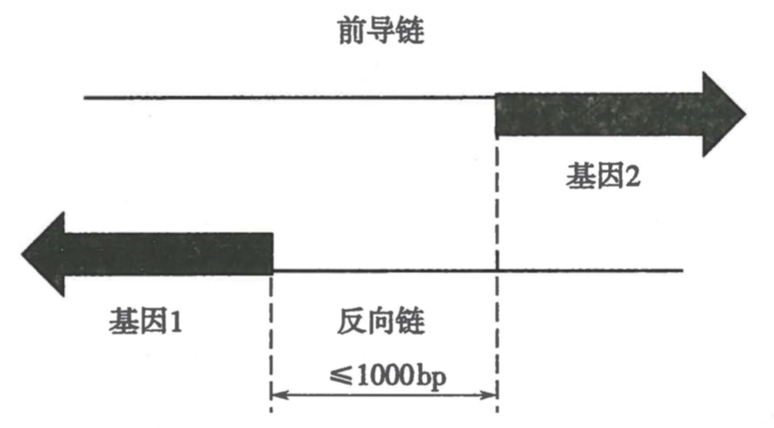
双向启动子
二、顺式作用元件和转录因子在真核转录起始中有重要作用
-
转录起始相关的顺式作用元件
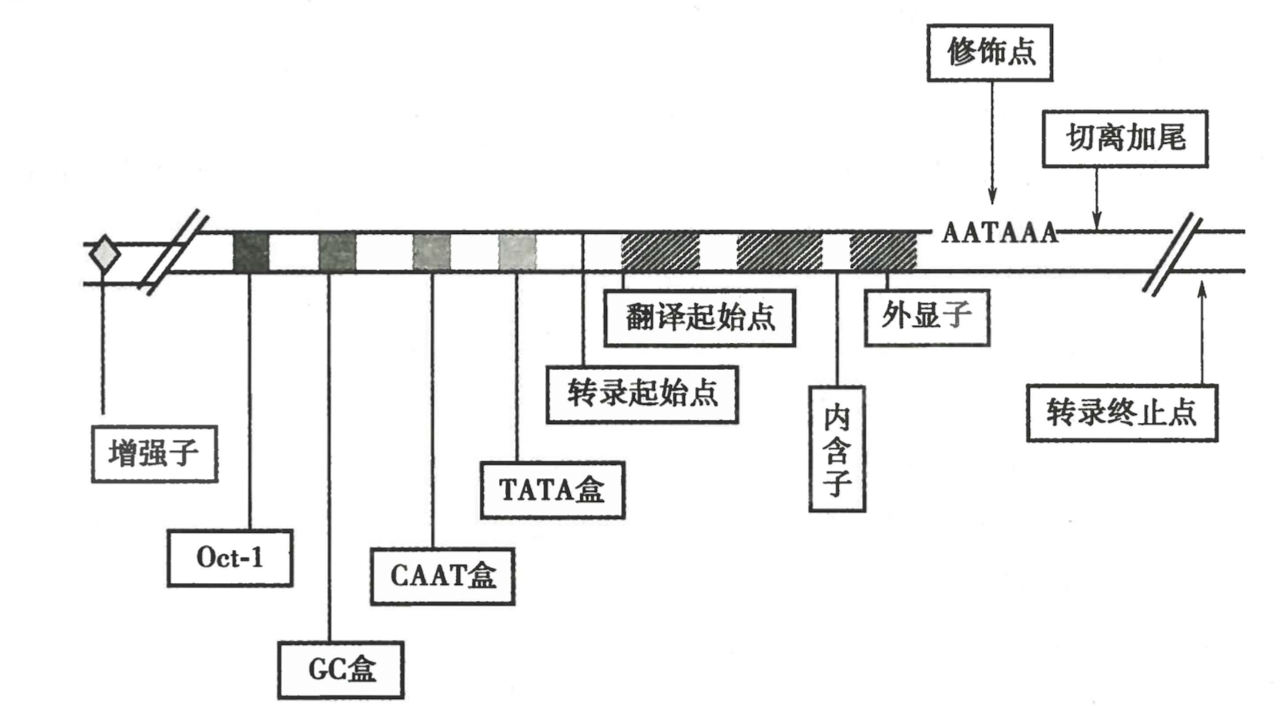
- 核心启动子序列
- TATA box(Hognest box)
- 某些 house-keeping gene 没有 TATA box
- 起始子(initiator,Inr)
- 位于转录起始位点附近
- Pol II 保守识别
- TATA box(Hognest box)
- 启动子上游元件(近端序列)
- CAAT box 和 GC box
- 上游 40~200bp
- 结合 TF,增强转录效率
- CAAT box 和 GC box
- 增强子(远端序列)
- 1000bp-50000bp
- 可以调控上下游的启动子
- 核心启动子序列
-
转录因子 TF:
- 分类
-
TF I
-
TF II

-
TFII D:
- TBP(TATA box-binding protein):保证基础转录
- TAF(TBP-associated factor):Co-activator 辅激活因子,人体中 12 种,结合不同启动子
-
II中的四类转录因子
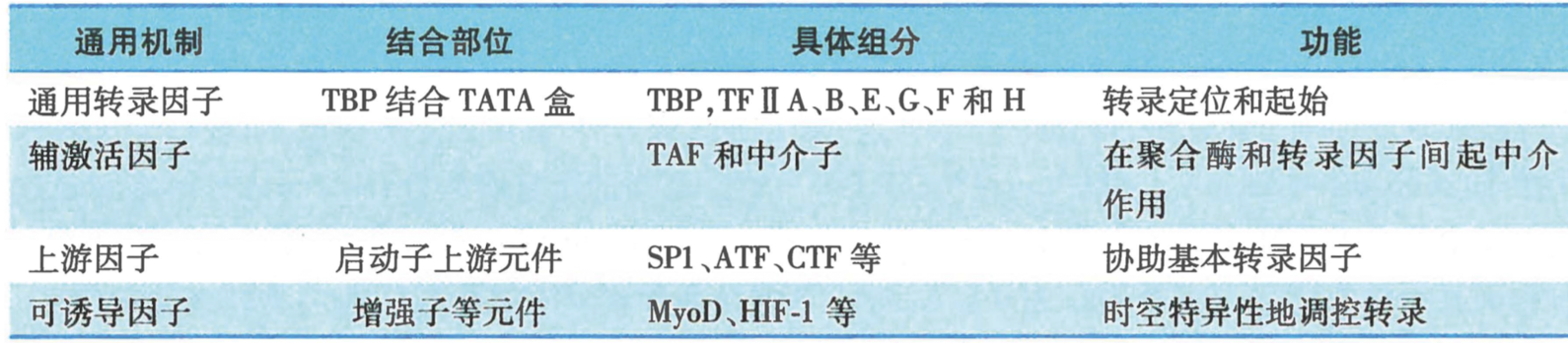
-
-
TF III
-
- 分类
-
转录起始前复合物
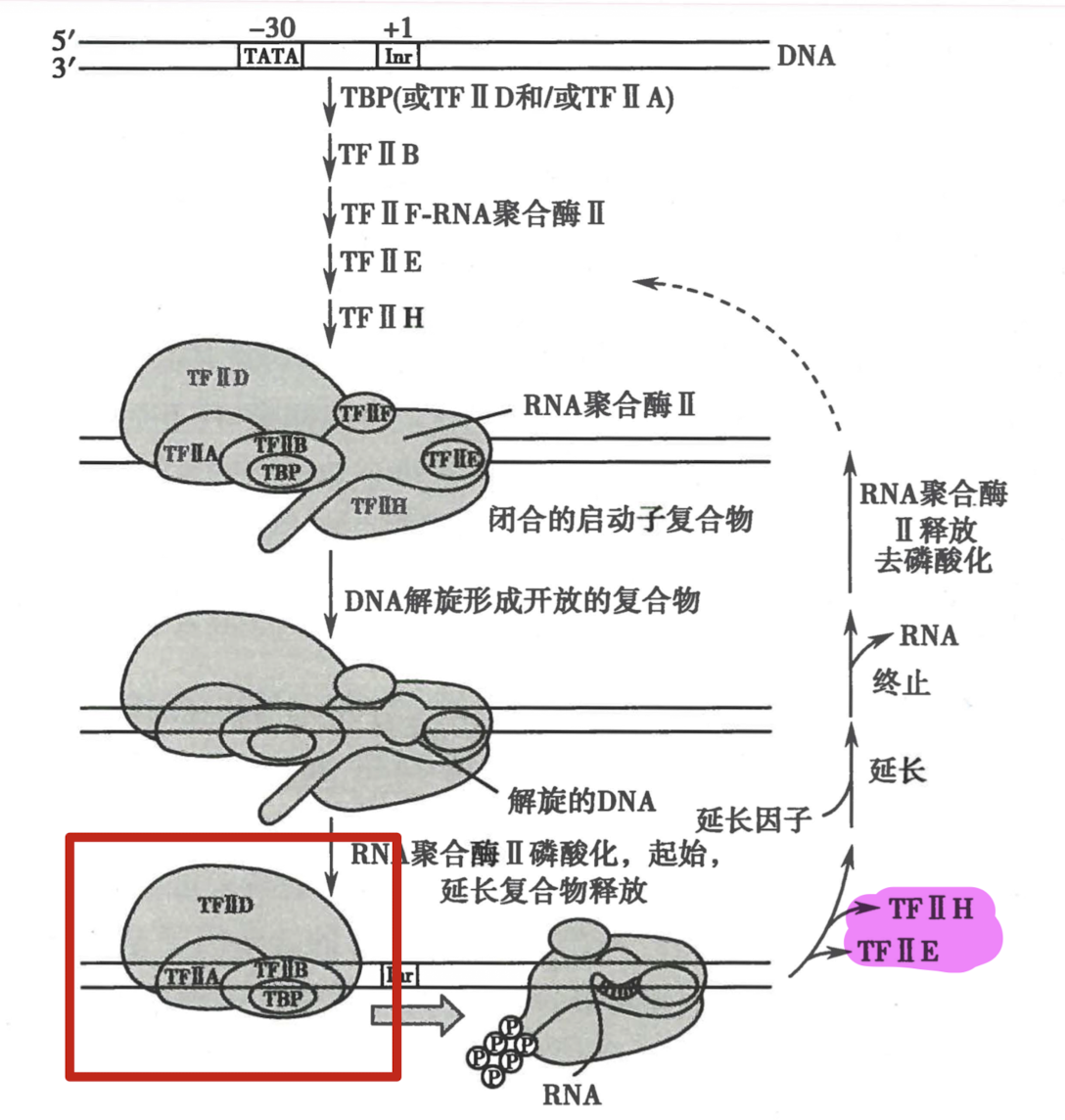
- TF II F:防止 pol II 与 DNA 非特异结合,协助 RNA pol II 与 启动子 (promoter)的靶向结合,延长阶段仍然结合 pol II
- TF II H:
- 解旋酶活性:解旋起始点附近的 DNA
- 激酶活性:磷酸化 CTD,改变开放复合体构象
-
少数反式作用因子的搭配启动特定基因转录
三、真核生物 RNA 转录延长过程与翻译不同步
- 转录过程中核小体只是发生位移
- 核小体的组蛋白富含精氨酸,精氨酸带正电与磷酸酸根负电荷作用,转录中精氨酸被乙酰化
四、真核生物的转录终止和加尾修饰同时进行
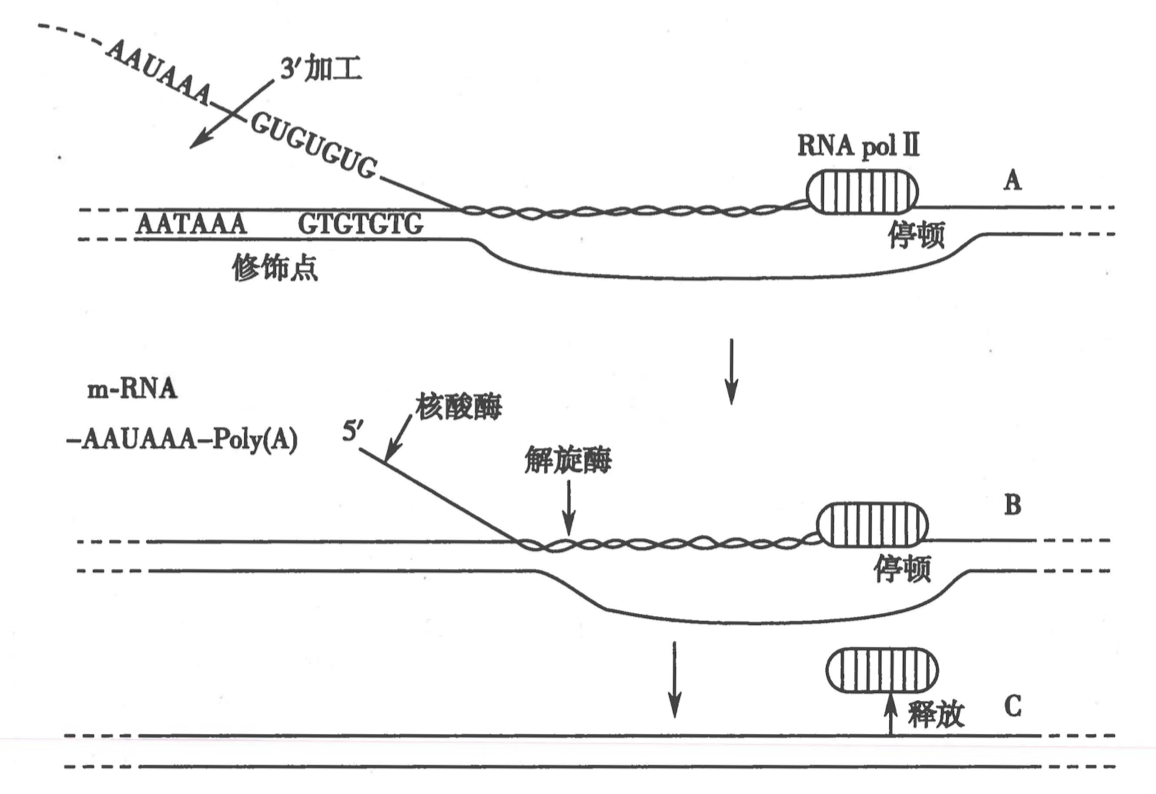
- 转录不是在 poly A 位置(AATAAA)停止,而是超出后停止
- 前体 mRNA 在修十点被切断,假如 polyA tail 和 5’ cap,下游虽然转录但是很快被降解
- RNA pol 没有3’-5’ exonuclease 功能,转录有一定错误率
第四节 真核生物前体 RNA 的加工和降解
一、前体 mRNA 经过首、尾修饰、剪接和编辑加工后才能成熟
- 5‘ cap:
-
加帽时间:25~30nt 时期
-
结构:7-甲基鸟嘌呤-ppp-5‘RNA
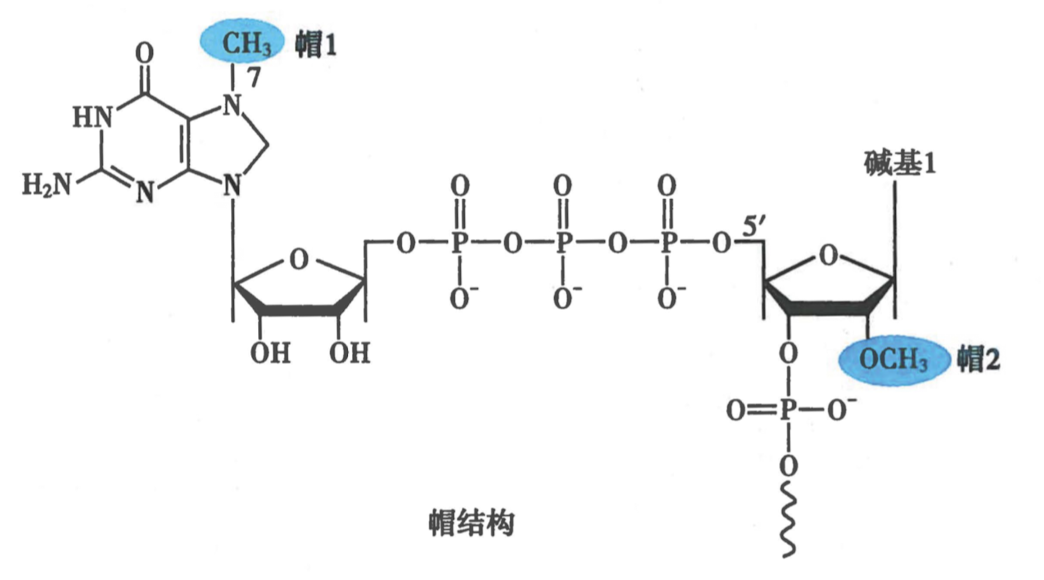
-
机理:
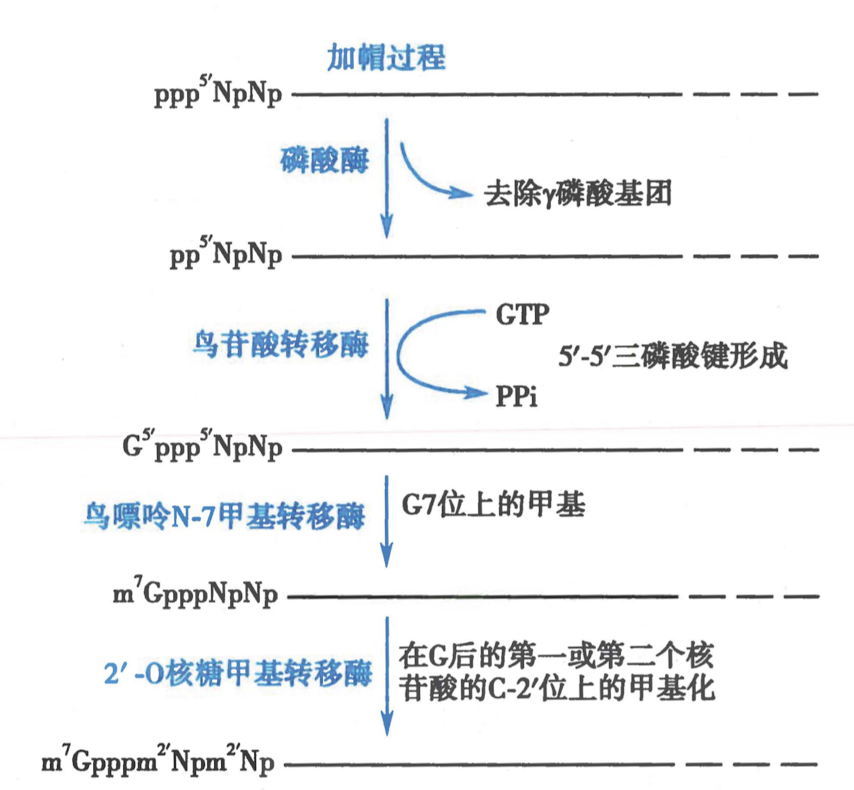
- 加帽酶:与 RNA pol II 的 CTD 结合,具有磷酸酶活性
- 甲基转移酶
-
作用:
-
使 mRNA 免遭核酸酶攻击
-
与帽结合蛋白质复合体结合,参与 mRNA 和核糖体的结合
-
-
- 前体 mRNA 在 3’特意位点断裂加上 polyA
- 长度:80-250nt
- 特点:
- polyA 不依赖 DNA 模板
- pre-mRNA 长度大于成熟 mRNA
- 核酸内切酶切割
- 尾部修饰和转录终止同时进行
- poly A的长度与 mRNA 的寿命呈现正相关
- 特例:组蛋白 gene 没有 poly A
- 前体 mRNA 的剪接主要是去除内含子
-
内含子总成套索 RNA 被剪除(lariat RNA)
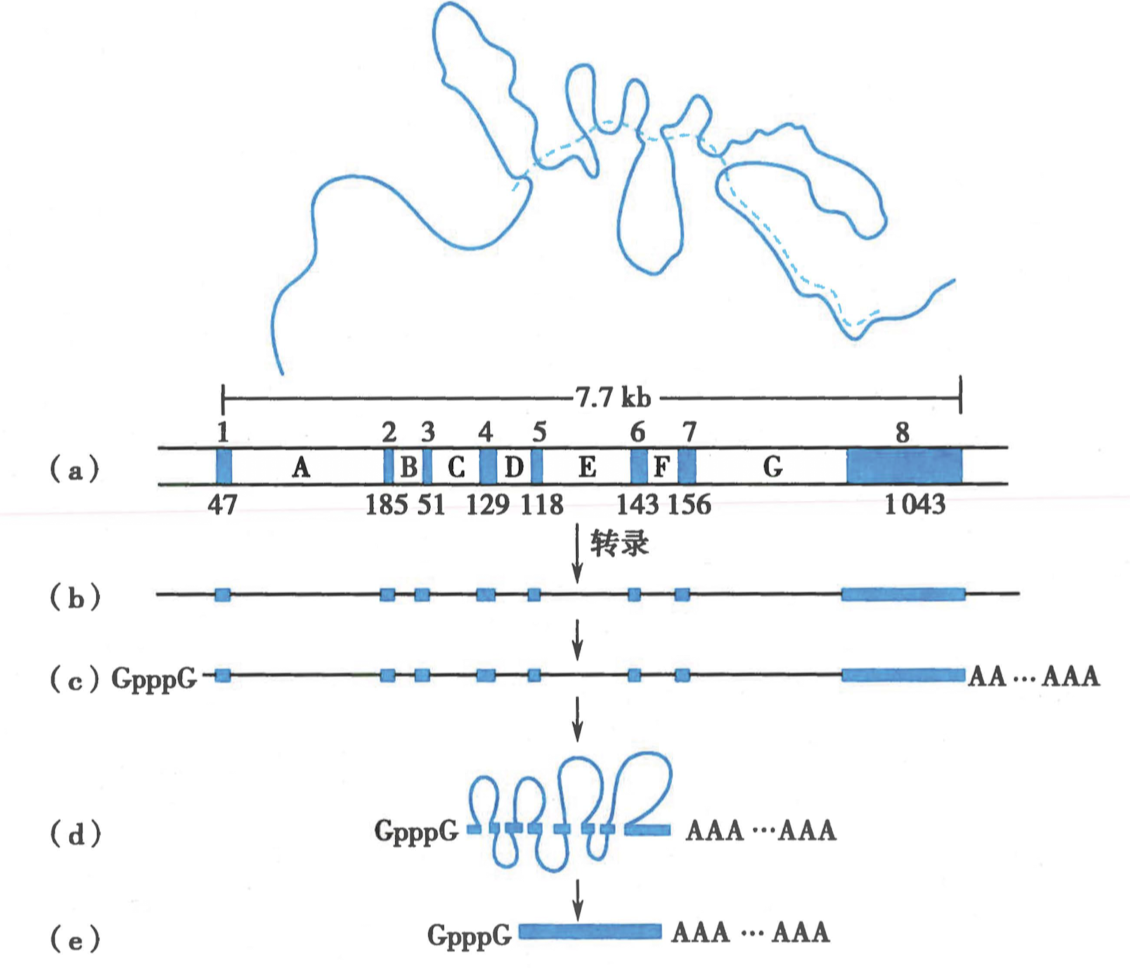
-
内含子在剪接接口处剪除
- pre-mRNA 含有可以被剪接体识别的特殊保守序列
- 5‘-GU ······ AG-OH-3’(剪接接口):5‘-剪接位点,剪接分支点,3’-剪接位点
-
剪接过程需要两次转酯反应(不耗能)
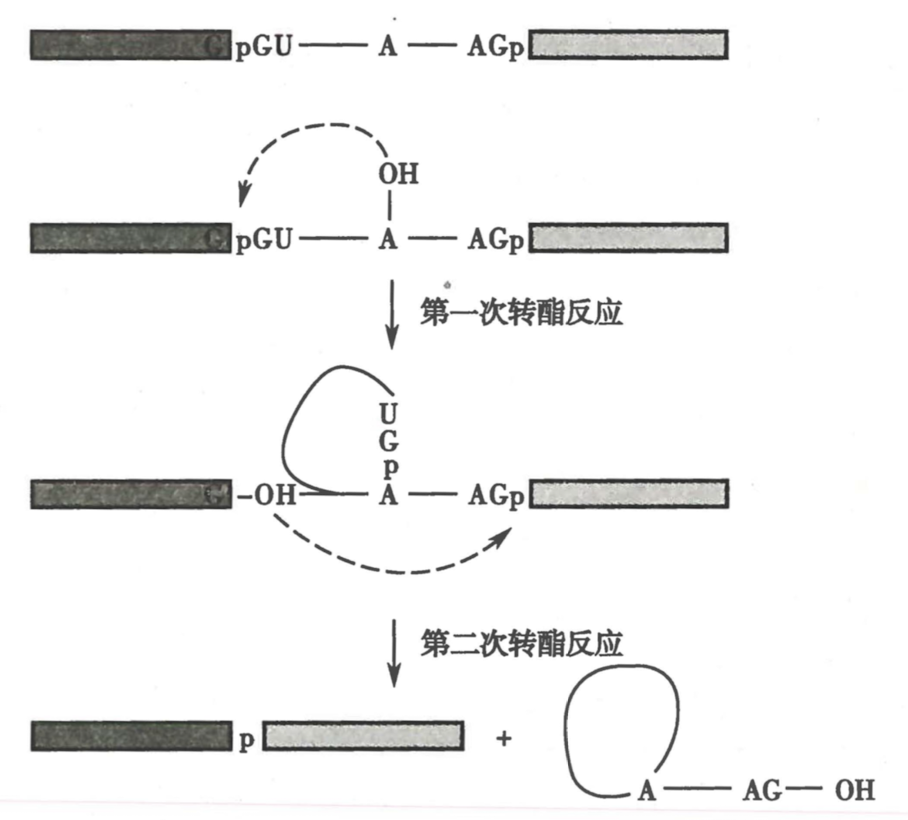
-
剪接体(Splicesome)是内含子剪接场所
- 组成:
- 5 种 snRNA(U1,U2…富含尿嘧啶) 和 100 种以上 protein
- 每一种 snRNA分别于多种 pro 结合 5 种 snRNP
- 金属离子 U6 催化剪接反应
- 组成:
-
前体 mRNA 分子有剪切和剪接两种模式
-
前体 mRNA 分子可以发生可变剪接(alternative splicing)
-
- mRNA 编辑是对基因的编码序列进行转录后加工
- 分化加工:mRNA 的 nt 位可以被改变
二、真核前体 rRNA 经过剪切形成不同类别的 rRNA
-
rRNA gene 属于冗余基因(redundant gene)
- 定义:染色体上相似或完全一样的纵裂串联基因(randem gene)单位的重复。
- 举例:5S rRNA,组蛋白 gene,免疫球蛋白 gene
- 每个 gene 被不能转录的基因间隔(gene spacer)分段分开
-
18S,5.8S,28S rRNA gene 串联在一起
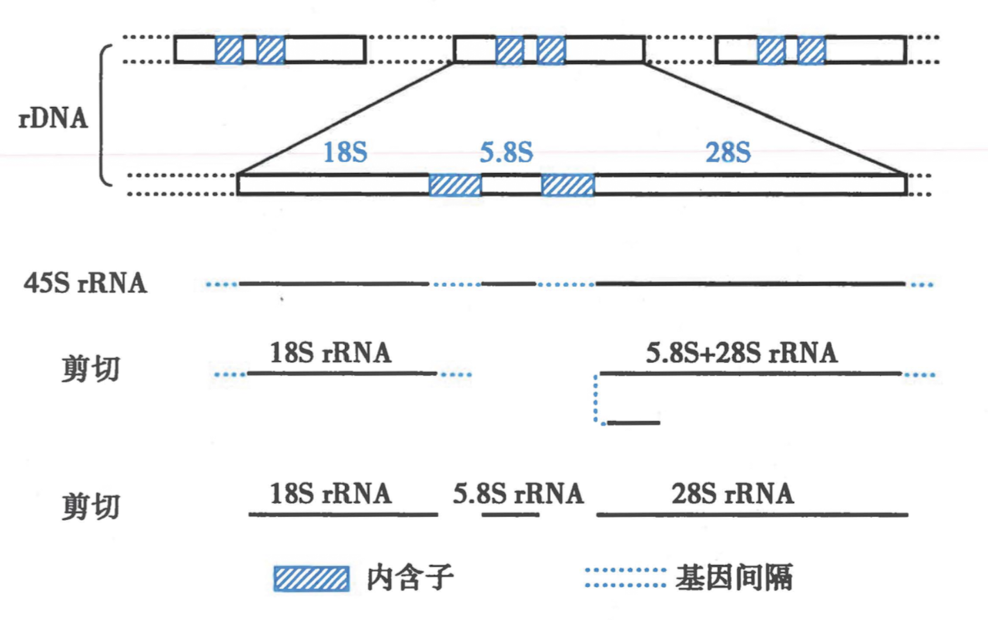
三、真核前体 tRNA 的加工包括核苷酸的碱基修饰
- 真核生物有 40-50 中不同的 tRNA
- 加工过程:
-
5‘ 核苷酸前导序列由 RNase P 切除
RNase P为核酶(Ribozyme) -
3’的两个 U 被 RNase Z 切除
-
核苷酸转移酶添加 CCA
-
碱基化学修饰
-
tRNA 剪切内切酶(TSEN)切除内含子,tRNA 连接酶催化连接
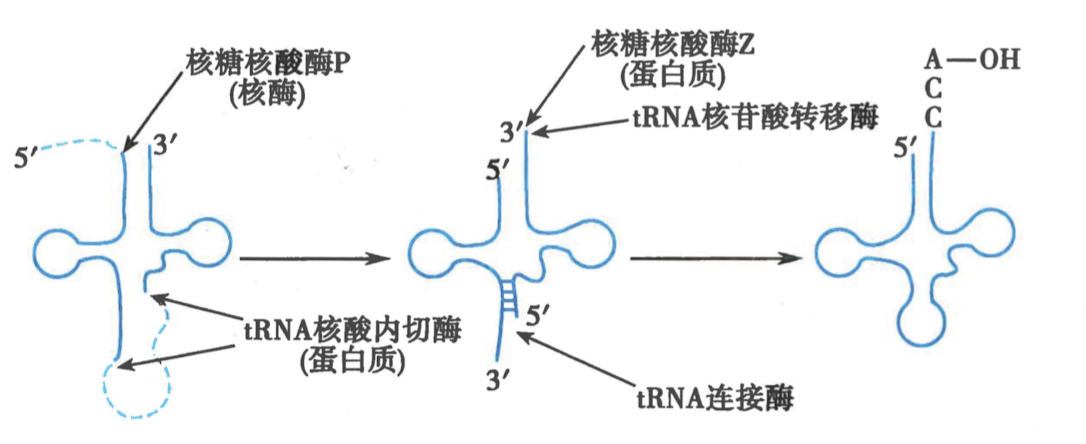
-
四、RNA 催化一些内含子的自剪接(Self-splicing)
- 定义:RNA 分子催化自身内含子剪接的反应
五、真核 RNA 在细胞内的降解有多种途径
-
依赖脱腺苷酸化的 mRNA 降解是重要的 mRNA 代谢途径
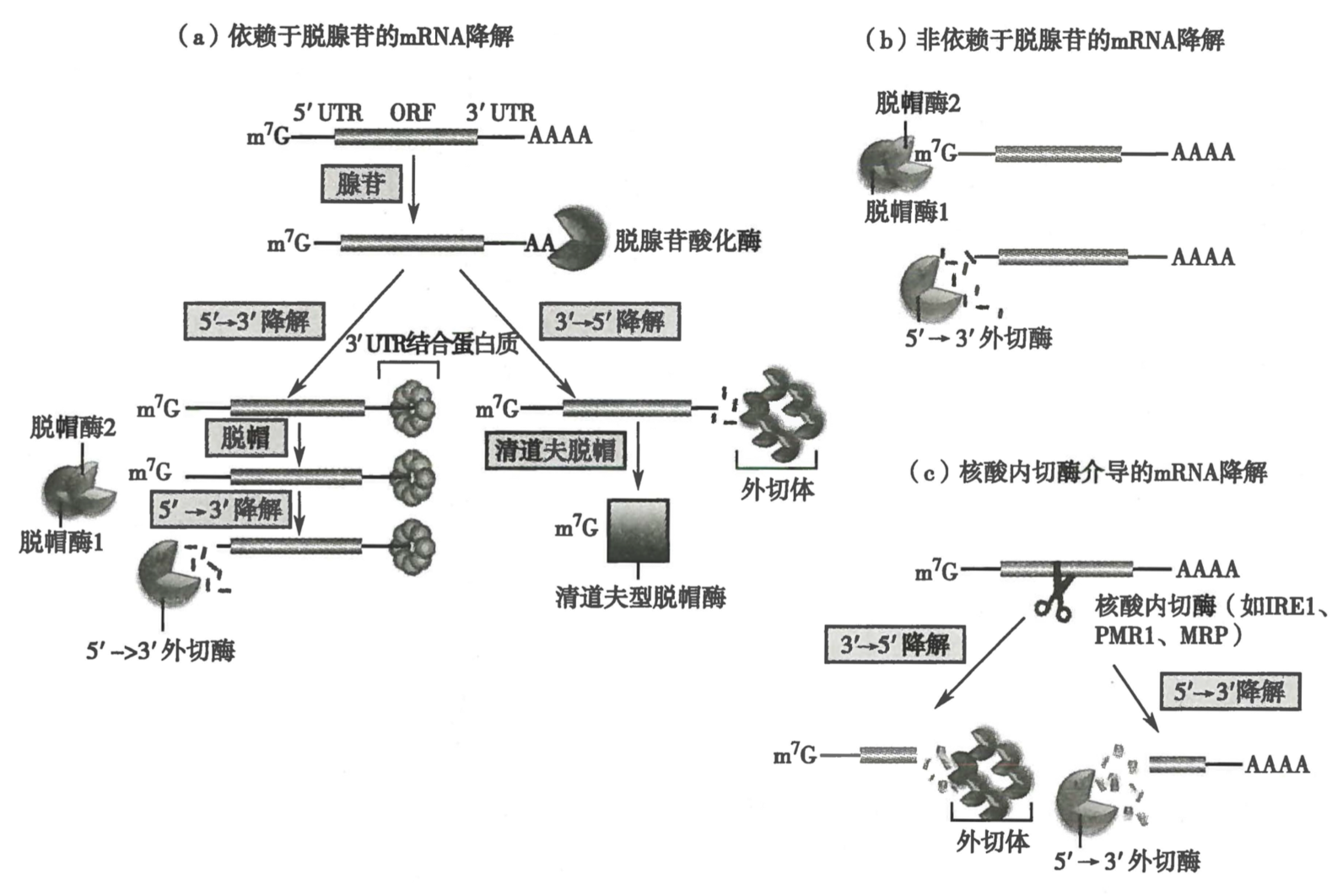
- 途径 1
- mRNA 形成封闭环状结构以防止脱腺苷酸化酶和脱帽酶的攻击
- 脱腺苷酸化酶:入侵环状结构,消除 3‘ polyA tail
- 脱帽酶:结合 mRNA 的 5’端,对 7-甲基鸟嘌呤水解
- 途径 2:少部分 mRNA 可以不被脱腺苷酸化酶处理直接脱帽
- 途径 3:有些 mRNA 可以被核酸内切酶切割
- 途径 1
-
无义介导的 mRNA 降解是一种重要的真核 mRNA 质量监控机制
__EOF__

本文链接:https://www.cnblogs.com/Fang-Hao/p/18369203.html
关于博主:评论和私信会在第一时间回复。或者直接私信我。
版权声明:本博客所有文章除特别声明外,均采用 BY-NC-SA 许可协议。转载请注明出处!
声援博主:如果您觉得文章对您有帮助,可以点击文章右下角【推荐】一下。您的鼓励是博主的最大动力!


【推荐】国内首个AI IDE,深度理解中文开发场景,立即下载体验Trae
【推荐】编程新体验,更懂你的AI,立即体验豆包MarsCode编程助手
【推荐】抖音旗下AI助手豆包,你的智能百科全书,全免费不限次数
【推荐】轻量又高性能的 SSH 工具 IShell:AI 加持,快人一步
· 被坑几百块钱后,我竟然真的恢复了删除的微信聊天记录!
· 没有Manus邀请码?试试免邀请码的MGX或者开源的OpenManus吧
· 【自荐】一款简洁、开源的在线白板工具 Drawnix
· 园子的第一款AI主题卫衣上架——"HELLO! HOW CAN I ASSIST YOU TODAY
· Docker 太简单,K8s 太复杂?w7panel 让容器管理更轻松!