
易基因:上海交大王琼团队揭示TGFβ-Smad信号通路和DNA甲基化在调控胚胎干细胞命运中的作用|NC
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
TGF-β超家族包括TGFβ、Activin、Nodal、BMPs等,通过受体丝氨酸/苏氨酸激酶传递信号,对细胞生长、分化、组织再生和癌变至关重要。Smad2和Smad3(Smad2/3)在TGFβ激活后被磷酸化,形成三聚体,可以与共同介质Smad4结合或不结合。Smad4与Smad2和Smad3(Smad2/3)结合形成转录调控复合体在典型的TGFβ信号通路中至关重要,但并非所有响应都必需,不依赖Smad4为共同介质的Smad2/3作用仍研究不足。因此研究Smad2/3在独立于Smad4情况下如何调控mESCs谱系起始和分化,并探索这一过程中的分子机制尤为重要。
近日,上海交通大学基础医学院王琼研究员、Yanhua Du和生命科学技术学院章永春团队合作,共同探讨了TGFβ-Smad信号通路和DNA甲基化在调控小鼠胚胎干细胞(mESCs)从原始态多能性(naïve pluripotency)向始发态多能性(primed pluripotency)转变以及随后的分化过程中的作用。相关研究成果以“A stepwise mode of TGFβ-SMAD signaling and DNA methylation regulates naïve-to-primed pluripotency and differentiation”为题发表在Nature子刊《Nature Communications》上。

标题:A stepwise mode of TGFβ-SMAD signaling and DNA methylation regulates naïve-to-primed pluripotency and differentiation(TGFβ-SMAD信号通路和DNA甲基化模式调控原始态到始发态多能性转换和分化)
期刊:Nature Communications
影响因子:IF 14.7
应用技术:RNA-seq、ChIP-seq、CUT&Tag、ATAC-seq(易基因金牌技术)
本研究引入了一个逐步模式,其中Smad2/3通过与不同效应因子合作来调控小鼠胚胎干细胞(mESCs)的谱系启动和分化。在从原始态多能性向始发态多能性转变的过程中,Smad2/3上调DNA甲基转移酶3b(Dnmt3b),建立了适当的DNA甲基化模式,进而使Smad2/3能够结合到外胚层标记基因的启动子和增强子的低甲基化区域。因此,在Smad2/3缺失情况下,Smad4单独不能启动外胚层特异性基因转录。当始发态外胚层细胞开始分化时,Dnmt3b在全基因组甲基化中的活跃度降低,Smad4与Smad2/3形成复合物以支持中胚层和内胚层诱导。因此,缺乏Smad4的mESCs可以经历启动过程,但在下游分化中遇到困难。本研究揭示了TGFβ信号及其在细胞过程中作用的复杂机制。
研究方法
- 使用CRISPR/Cas9编辑技术生成Smad2/3和Smad4敲除(KO)小鼠胚胎干细胞系。
- 通过RNA测序(RNA-seq)分析野生型(WT)、Smad2/3敲除(S2/3DKO)和Smad4敲除(S4KO)细胞的差异表达基因(DEGs),以及主成分分析(PCA)、基因本体(GO)分析、基因集变异分析(GSVA)。
- 利用染色质免疫沉淀测序(ChIP-seq)和CUT&Tag技术研究Smad2/3和Dnmt3b的基因组结合位点。
- ATAC-seq分析开放染色质区域,以了解染色质可及性。
- WGBS和MeDIP分析ES和EpiLC阶段WT和Dnmt3b敲除(3bKO)细胞的DNA甲基化水平。
- 免疫共沉淀(IP)和质谱(MS)用于鉴定与Smad2/3互作的蛋白质,包括Dnmt3b。
- Western blot检测特定蛋白质的表达水平;免疫荧光检测特定蛋白质在细胞内的表达和定位。
- 体外蛋白质-蛋白质互作实验:使用GST融合蛋白和His标签融合蛋白进行体外拉下实验,以研究蛋白质间的直接相互作用。
结果图形
(1)S2/3DKO 和 S4KO的的差异转录组分析
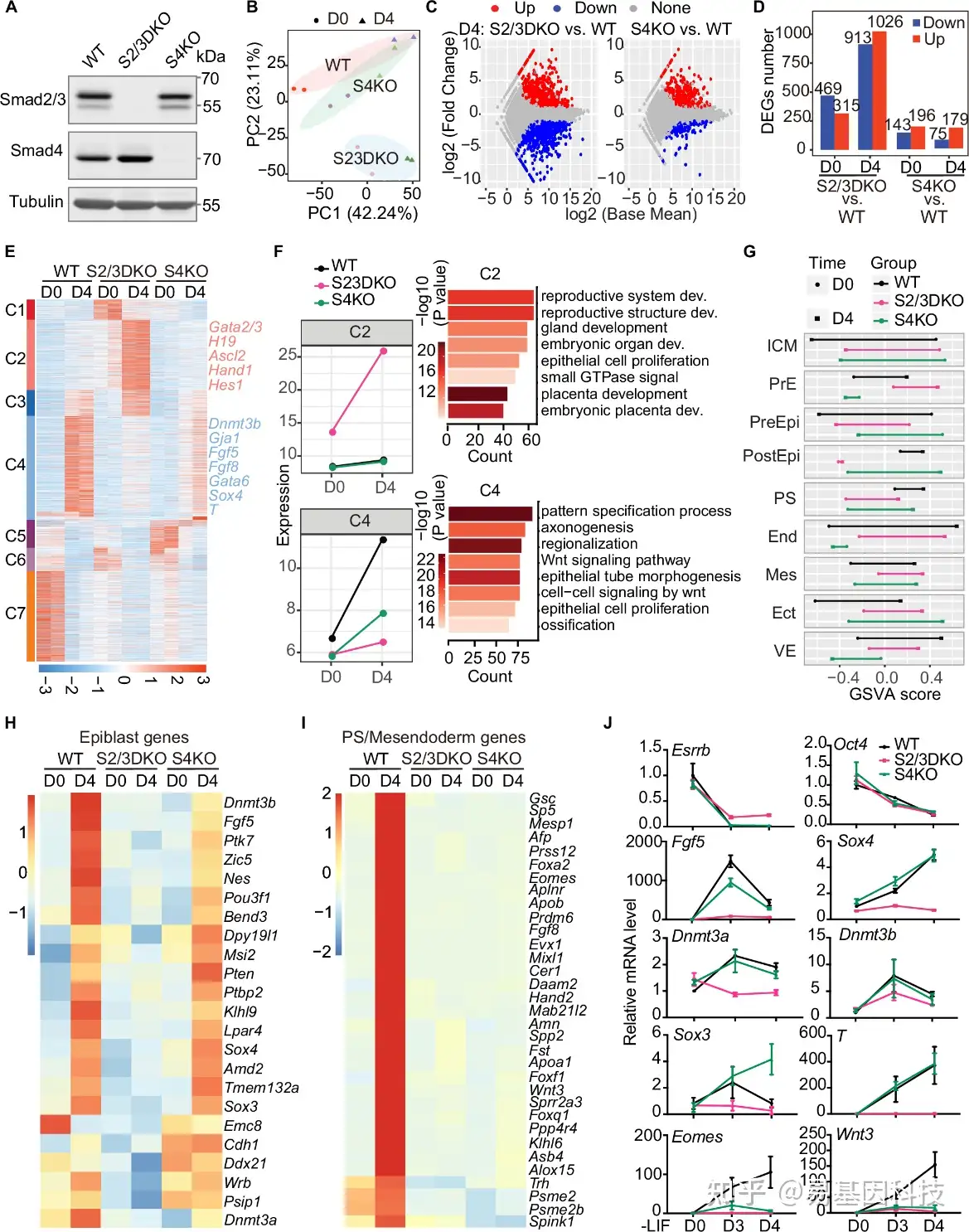
图1:mESC分化过程中Smad2/3和Smad4的差异需求。
- 对野生型(WT)、Smad2/3双敲除(S2/3DKO)和Smad4敲除(S4KO)细胞系中Smad2/3和Smad4蛋白的Western blot分析。Tubulin为对照。
- 对WT、S2/3DKO和S4KO在mESC(第0天,D0)和胚胎体(EB)分化4天(D4)产生的RNA-seq数据进行主成分分析(PCA)。
- WT、S2/3DKO或S4KO的D4 EBs的RNA-seq数据MA图。与WT相比,在S2/3DKO或S4KO中上调(红色)或下调(蓝色)基因。每个条件分析两个生物学重复样本。
- 分别以红色和蓝色条表示上调和下调的差异表达基因(DEGs)。
- 在D0或D4 EBs中WT、S2/3DKO或S4KO的所有DEGs热图。
- 左侧点线图显示(E)中C2和C4的平均表达模式。右侧显示C2和C4中基因的GO分析。
- 使用GSVA分析从D0到D4在WT、S2/3DKO和S4KO中的RNA-seq数据的谱系基因表达模式。ICM,内细胞团;PrE,原始内胚层;PreEpi,前外胚层;PostEpi,后外胚层;PS,原条;End,内胚层;Mes,中胚层;Ect,外胚层;VE,内脏内胚层。
- 热图显示WT、S2/3DKO和S4KO的mESC(D0)和D4 EBs中与外胚层相关的基因。
- 热图显示WT、S2/3DKO和S4KO的mESC(D0)和D4 EBs中的PS或中胚层基因。
- WT、S2/3DKO和S4KO的mESC(D0)、EB D3和D4中的谱系标记基因表达qRT-PCR分析。
(2)在Smad4缺失情况下,外胚层(Epiblast)可以形成
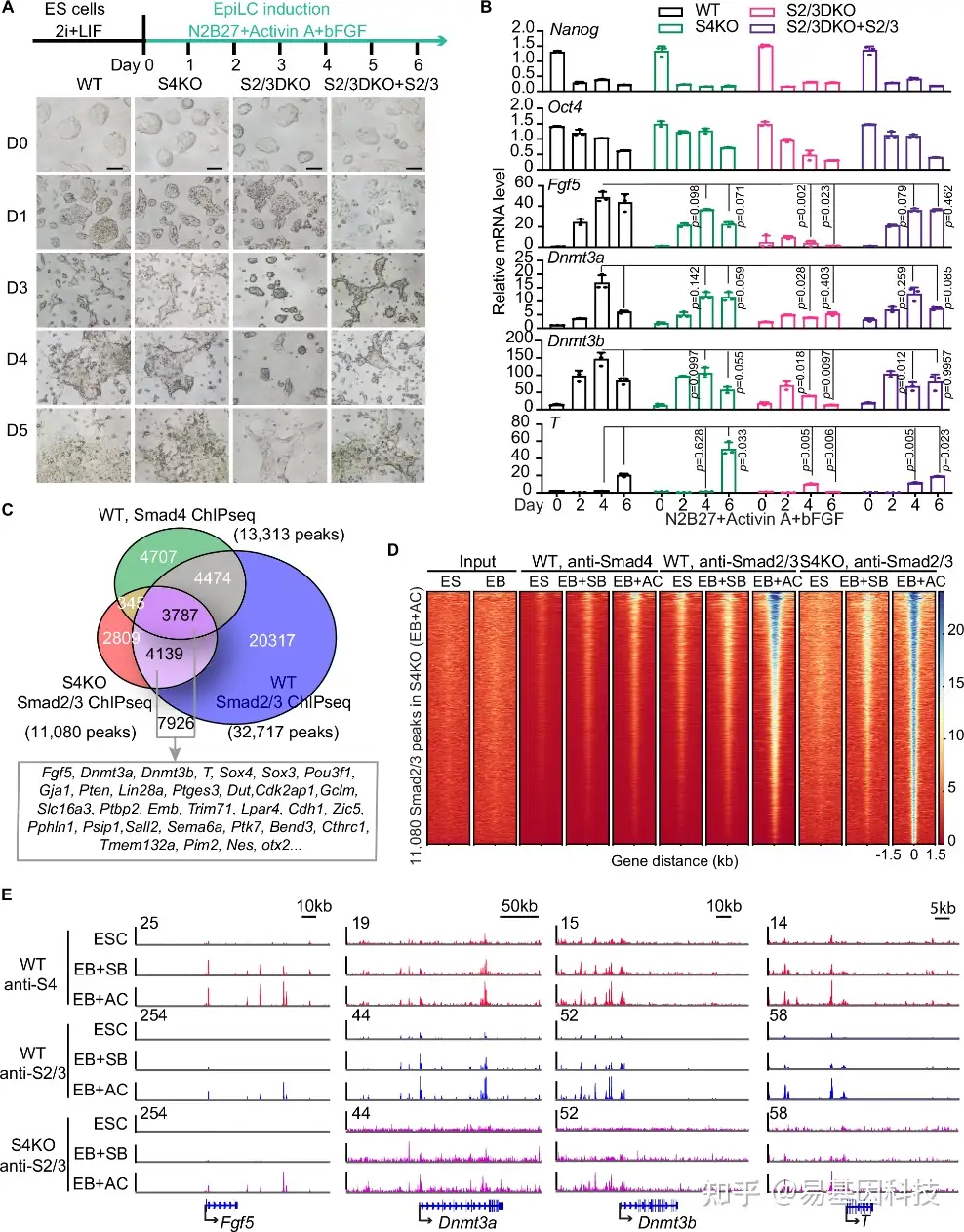
图2:Smad4缺失情况下的外胚层形成。
- 图表展示从mESC向EpiLC转变过程(顶部)。mESCs在从2i+LIF培养基转移到含有Activin A和bFGF的N2B27培养基中培养6天后,转变为EpiLCs。实验5次重复,展示WT、S4KO、S2/3DKO以及用Smad2/3拯救的S2/3DKO在转变过程中细胞形态变化的代表性图像(底部)。
- 通过qRT-PCR分析诱导EpiLCs期间多能性基因(Nanog, Oct4)和着床后外胚层基因(Fgf5, Dnmt3a/3b, T)的mRNA水平。
- 维恩图显示WT中Smad4 ChIP-seq、WT中Smad2/3 ChIP-seq和S4KO中Smad2/3 ChIP-seq之间的重叠峰值。注释了与7926个峰值相邻的基因,其中标记了始发态外胚层基因。
- 热图显示在AC处理的S4KO D3 EBs中,Smad2/3和Smad4的ChIP-seq标签密度在11080个高置信度Smad2/3结合位点±1.5 kb基因组区域内。SB,SB431542,TGFβR的选择性抑制剂;AC,Activin A,Nodal/Activin受体的可用配体。
- Fgf5、Dnmt3a、Dnmt3b和T位点的基因轨迹视图。在SB或AC处理的D3 EBs和未处理的D0 mESCs中进行Smad2/3和Smad4 ChIP-seq。底部示意性表示RefSeq基因结构。
(3)Dnmt3b与Smad2/3互作
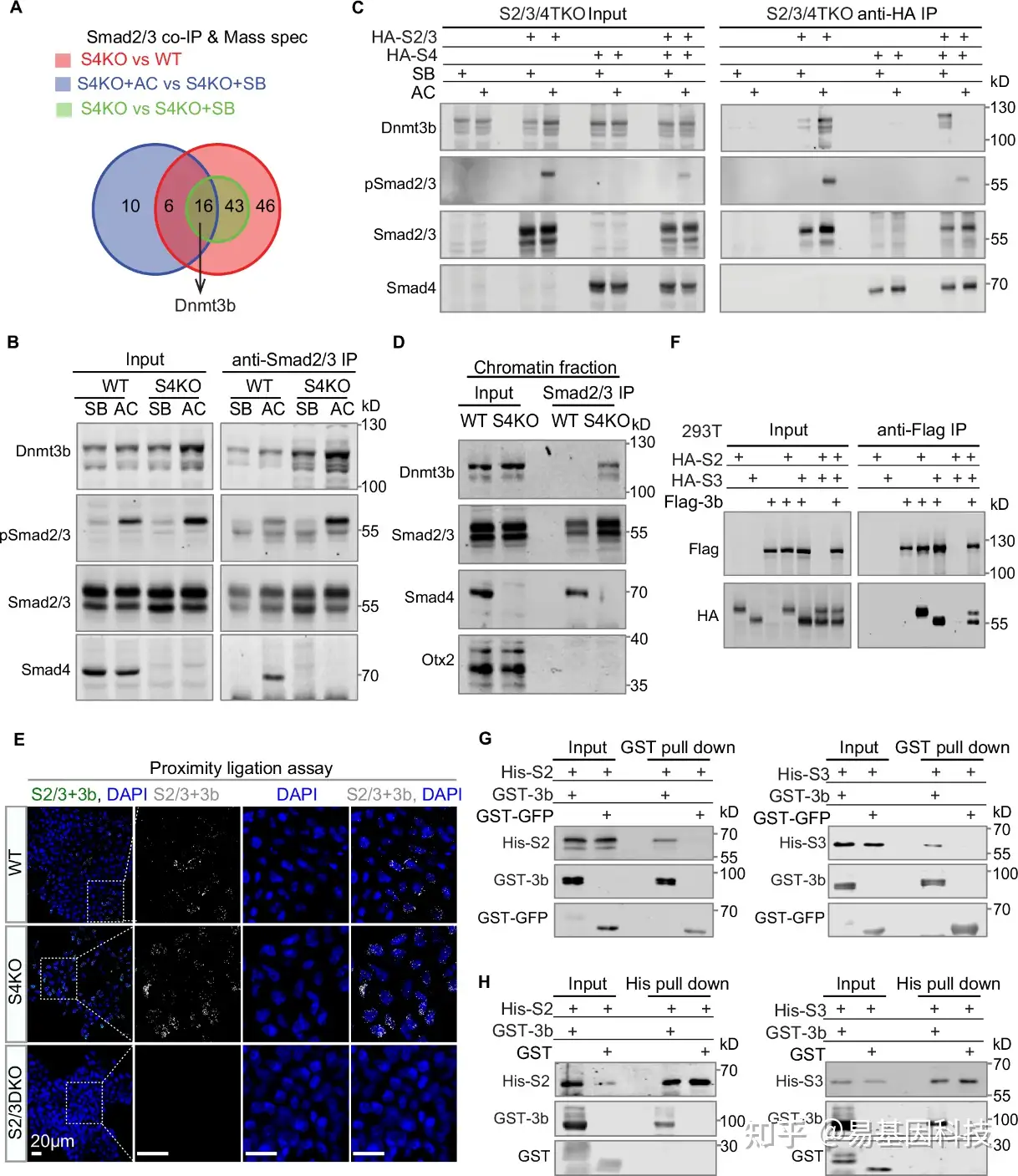
图3:Dnmt3b与Smad2/3互作。
- Smad2/3共免疫沉淀和质谱(IP-MS)分析维恩图。在WT和S4KO的D3 EBs中进行Smad2/3 IP-MS,包括未处理的、用SB或AC处理的样本。在所有三个比较中(S4KO与WT、S4KO+AC与S4KO+SB、S4KO与S4KO+SB)鉴定出16种蛋白质,包括Dnmt3b。
- 经SB或AC添加处理后,通过使用Dnmt3b、pSmad2/3、Smad2/3和Smad4抗体的western blot实验分析WT和S4KO D3 EBs中Smad2/3的免疫沉淀。
- 在Smad2/3/4TKO细胞中,HA标记的Smad2/3(HA-S2/3)或/和Smad4(HA-S4)被过表达,进行靶向HA的免疫沉淀western blot分析。细胞在SB或AC处理后收集。
- 在AC处理的WT和S4KO D3 EBs中,分析Smad2/3的染色质结合蛋白的免疫沉淀,使用了Dnmt3b、Smad2/3、Smad4和Otx2抗体的western blot分析。
- 展示WT、S4KO和S2/3DKO D3 EBs的PLA实验的代表性共聚焦图像。
- 在293T细胞中过表达HA标记的Smad2(HA-S2)或Smad3(HA-S3),或/和Flag标记的Dnmt3b(Flag-3b)。在进行抗Flag免疫沉淀后检测Flag和HA的western blot。
- 使用细菌表达的GST-Dnmt3b或GST-GFP,以及6x组氨酸标记的Smad2(His-S2,左)或Smad3(His-S3,右)进行GST拉下实验。
- 使用细菌表达的GST-Dnmt3b或GST,以及6x组氨酸标记的Smad2(His-S2,左)或Smad3(His-S3,右)进行His拉下实验。
(4)Smad2/3上调Dnmt3b对外胚层形成至关重要
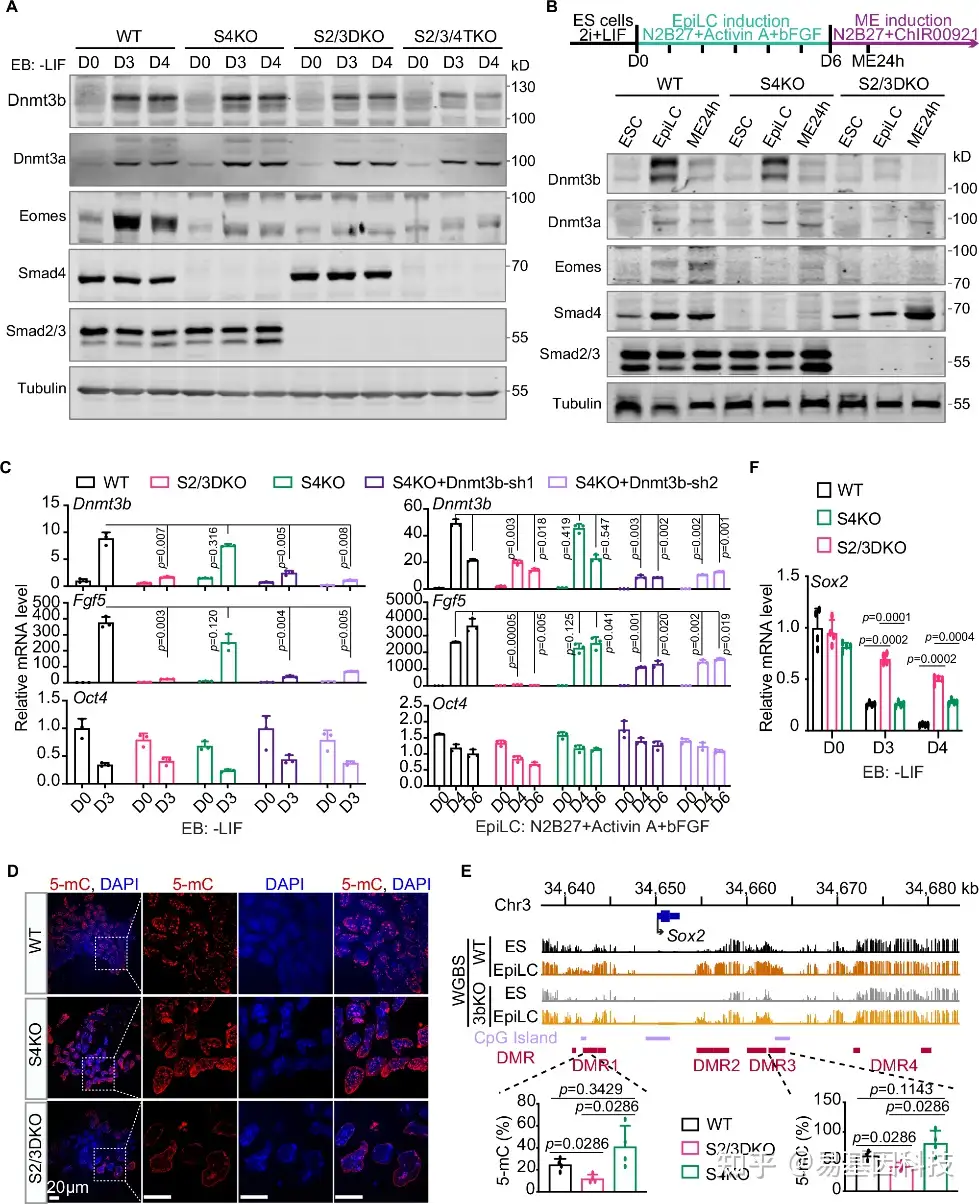
图4:Smad2/3上调Dnmt3b对外胚层形成非常重要。
- 对WT、S4KO、S2/3DKO和S2/3/4TKO细胞在EB分化(D0、D3和D4)期间的Dnmt3a、Dnmt3b、Eomes、Smad2/3和Smad4蛋白水平进行western blot分析。
- mESC-EpiLC-ME两步诱导方案(顶部)。将细胞转移到含有Activin A和bFGF的N2B27培养基中培养6天后,EpiLCs进一步在加入CHIR99021的N2B27培养基中培养24小时以形成ME。western blot显示WT、S4KO和S2/3DKO中Dnmt3a、Dnmt3b、Eomes、Smad4和Smad2/3的表达。
- 对WT、S2/3DKO、S4KO以及带有Dnmt3b shRNA 1或2的S4KO细胞的EB分化(D0、D3)和EpiLC诱导(D0、D4、D6)期间的始发态外胚层基因(Dnmt3b、Fgf5)和多能性基因Oct4进行qRT-PCR分析。
- WT、S4KO和S2/3DKO EBs中5-mC(红色)和DAPI(蓝色)染色的代表性共聚焦显微镜图像。
- IGV视图显示WT和Dnmt3b敲除(3bKO)中Sox2位点在ES和EpiLC阶段的WGBS信号谱。CpG岛(紫色)和差异甲基化区域(DMRs)(红色)也进行标记。在WT、S2/3DKO和S4KO的D3 EBs中进行Sox2 DMR1和DMR3的DNA甲基化水平的MeDIP-qPCR分析。
- qRT-PCR检测WT、S2/3DKO和S4KO细胞在EB分化前后(D0、D3、D4)的Sox2 mRNA水平。
(5)Dnmt3b独立于Smad4介导Smad2/3与外胚层基因结合。
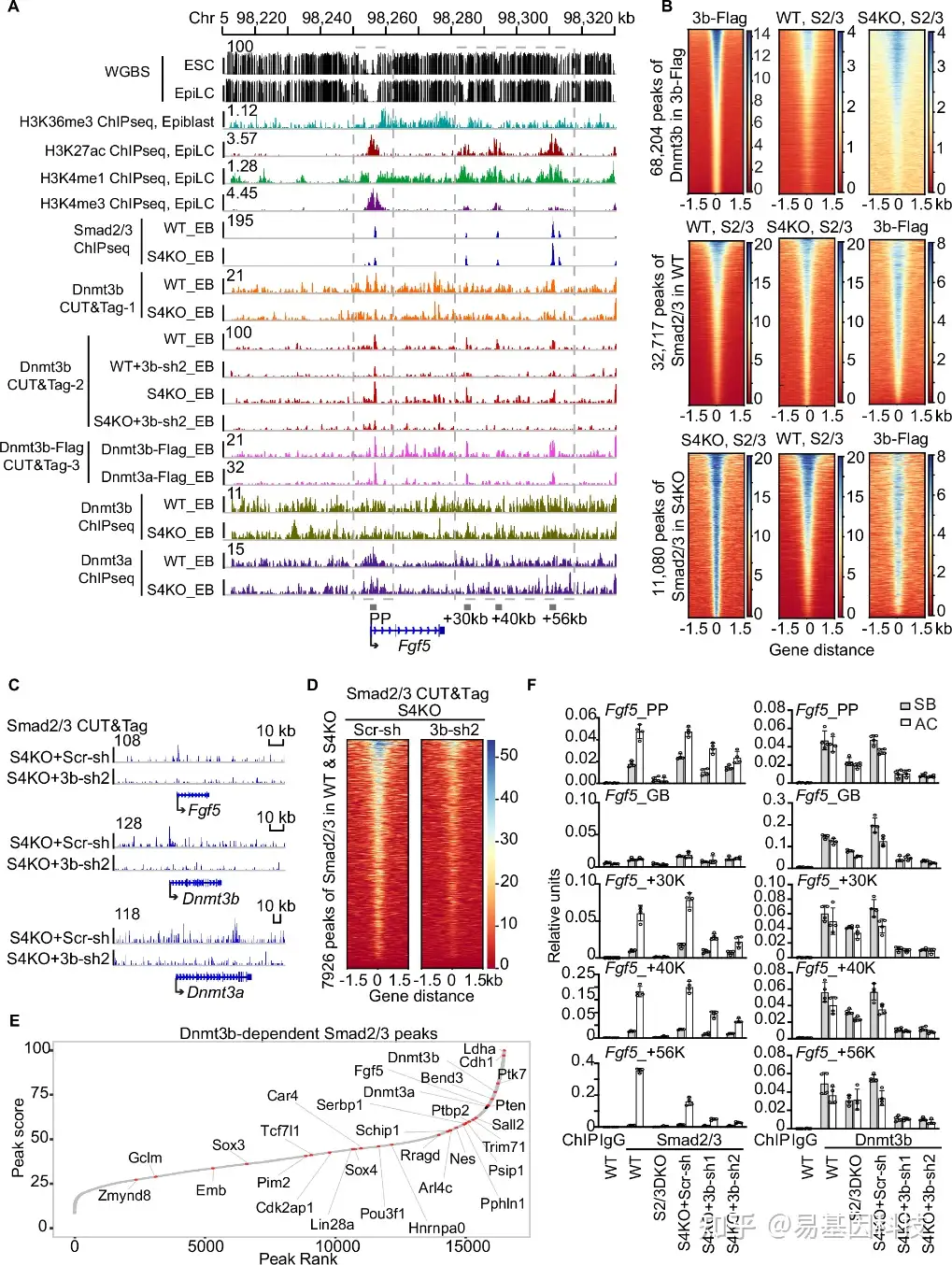
图5:Dnmt3b独立于Smad4介导Smad2/3与外胚层基因结合。
- Fgf5基因位点的WGBS、ChIP-seq和CUT&Tag分析综合视图。WGBS以及H3K27ac、H3K4me1和H3K4me3的ChIP-seq在ESC或EpiLC阶段进行,H3K36me3 ChIP-seq在小鼠E6.5外胚层进行。Smad2/3 ChIP-seq在WT和S4KO的EBs中进行(本研究),Dnmt3b CUT&Tag在WT、S4KO、WT+Dnmt3b-shRNA2(WT+3b-sh2)和S4KO+Dnmt3b-shRNA2(S4KO+3b-sh2)的EBs中进行,Flag CUT&Tag实验在Dnmt3a-Flag或Dnmt3b-Flag细胞中进行;WT和S4KO EBs中Dnmt3a或Dnmt3b的ChIP-seq显示在底部。
B 在Dnmt3b-Flag EBs中68204个Dnmt3b结合位点±1.5 kb基因组区域内Flag CUT&Tag的标签密度热图,以及在WT或S4KO中32717或11080个Smad2/3结合位点中心的Smad2/3 ChIP-seq。
C. S4KO和S4KO+3b-sh2的EBs中Smad2/3 CUT&Tag的基因轨迹视图,靶向Fgf5、Dnmt3a和Dnmt3b位点。
D. 在S4KO和S4KO+3bsh2周围7926个Smad4独立的Smad2/3峰值的Smad2/3 CUT&Tag信号热图。
E. Dnmt3b依赖的Smad2/3结合位点峰值得分图。标记了与峰值相邻的外胚层标记基因。
F. 对WT、S2/3KO、S4KO+Scr-shRNA、S4KO+3b-Sh1或S4KO+3b-sh2细胞系的D3 EBs中Fgf5启动子(Fgf5_PP)、基因体(Fgf5_GB)和增强子(Fgf5_+30k、Fgf5_+40k和Fgf5_+56k)的Smad2/3和Dnmt3b结合进行ChIP-qPCR分析。使用抗IgG的ChIP作为阴性对照。
易小结
本研究通过RNA-seq、ChIP-seq、CUT&Tag、ATAC-seq、WGBS等分析揭示了Smad2/3在mESCs向EpiLCs转变过程中通过诱导Dnmt3b表达来调控DNA甲基化模式,从而促进原始态多能性向始发态多能性的转变。同时还揭示了在Smad2/3缺失情况下,Smad4单独不能启动Epiblast特异性基因转录。在分化过程中,Dnmt3b活性降低,Smad4与Smad2/3形成复合物,支持中胚层和内胚层的诱导。
总之,本研究阐明了TGFβ信号通路在细胞过程中的复杂机制,特别是Smad2/3和Smad4在mESCs的原始态多能性向始发态多能性转变以及随后分化中的作用,提出了一个逐步调控模型。
关于易基因染色质免疫共沉淀测序 (ChIP-seq)
染色质免疫共沉淀(Chromatin Immunoprecipitation,ChIP),是研究体内蛋白质与DNA相互作用的经典方法。将ChIP与高通量测序技术相结合的ChIP-Seq技术,可在全基因组范围对特定蛋白的DNA结合位点进行高效而准确的筛选与鉴定,为研究的深入开展打下基础。
DNA与蛋白质的相互作用与基因的转录、染色质的空间构型和构象密切相关。运用组蛋白特定修饰的特异性抗体或DNA结合蛋白或转录因子特异性抗体富集与其结合的DNA片段,并进行纯化和文库构建,然后进行高通量测序,通过将获得的数据与参考基因组精确比对,研究人员可获得全基因组范围内某种修饰类型的特定组蛋白或转录因子与基因组DNA序列之间的关系,也可对多个样品进行差异比较。
应用方向:
ChIP 用来在空间上和时间上不同蛋白沿基因或基因组定位
- 转录因子和辅因子结合作用
- 复制因子和 DNA 修复蛋白
- 组蛋白修饰和变异组蛋白
技术优势:
- 物种范围广:细胞、动物组织、植物组织、细菌微生物多物种富集经验;
- 微量建库:只需5ng以上免疫沉淀后的DNA,即可展开测序分析;
- 方案灵活:根据不同的项目需求,选择不同的组蛋白修饰特异性抗体。
技术路线:
关于易基因全基因组重亚硫酸盐测序(WGBS)
全基因组重亚硫酸盐甲基化测序(WGBS)可以在全基因组范围内精确的检测所有单个胞嘧啶碱基(C碱基)的甲基化水平,是DNA甲基化研究的金标准。WGBS能为基因组DNA甲基化时空特异性修饰的研究提供重要技术支持,能广泛应用在个体发育、衰老和疾病等生命过程的机制研究中,也是各物种甲基化图谱研究的首选方法。
易基因全基因组甲基化测序技术通过T4-DNA连接酶,在超声波打断基因组DNA片段的两端连接接头序列,连接产物通过重亚硫酸盐处理将未甲基化修饰的胞嘧啶C转变为尿嘧啶U,进而通过接头序列介导的 PCR 技术将尿嘧啶U转变为胸腺嘧啶T。
应用方向:
WGBS广泛用于各种物种,要求全基因组扫描(不错过关键位点)
- 全基因组甲基化图谱课题
- 标志物筛选课题
- 小规模研究课题
技术优势:
- 应用范围广:适用于所有参考基因组已知物种的甲基化研究;
- 全基因组覆盖:最大限度地获取完整的全基因组甲基化信息,精确绘制甲基化图谱;
- 单碱基分辨率:可精确分析每一个C碱基的甲基化状态。

易基因染色质可及性-转座酶易接近染色质测序(ATAC-seq)
ATAC-seq(Assay for Transposase-Accessible Chromatin with high throughput sequencing)是通过使用高通量测序对转座酶可接近性核染色质区域进行分析的一种创新表观遗传学研究技术。该技术通过转座酶对某种特定时空下开放的核染色质区域进行切割,进而获得在该特定时空下基因组中所有活跃转录的调控序列。
真核生物的DNA并不是裸露的,而是被包装成核小体形成串珠状结构并进一步被折叠、包装。而基因的转录,需要将这种高级结构解开,使DNA成为可以使各种转录机器与其结合的裸露状态,即形成开放染色质区域。如何鉴定开放染色质区域呢?传统的方法主要是借助和DNase-Seq、MNase-Seq及ChIP-seq。但这些方法需要的起始细胞量较大,对于少量样本和珍稀样本可行性不高。ATAC-Seq是一种新型的研究开放染色质的技术,利用Tn5转座酶进入并切割裸露的DNA,并同时连接上特异性的测序接头。因为切割和加接头一步完成,因此该技术可大大降低所需细胞起始量。
技术优势:
(1)所需细胞起始量低。
(2)应用范围广,适用于大部分物种及细胞类型。
实验策略:

信息分析:
易基因提供全面的表观基因组学(DNA甲基化、DNA羟甲基化)和表观转录组学(m6A、m5C、m1A、m7G、ac4C)、染色质结构与功能组学技术方案(ChIP-seq、ATAC-seq),详询易基因:0755-28317900。

参考文献:
Zhao B, Yu X, Shi J, Ma S, Li S, Shi H, Xia S, Ye Y, Zhang Y, Du Y, Wang Q. A stepwise mode of TGFβ-SMAD signaling and DNA methylation regulates naïve-to-primed pluripotency and differentiation. Nat Commun. 2024 Nov 22;15(1):10123. pii: 10.1038/s41467-024-54433-5. doi: 10.1038/s41467-024-54433-5. PubMed PMID: 39578449.
相关阅读:
项目文章 | WGBS+RNA-seq揭示松材线虫JIII阶段形成过程中的DNA甲基化差异
项目文章 | WGBS+RNA-seq揭示黄瓜作物的“源-库”关系受DNA甲基化调控
 浙公网安备 33010602011771号
浙公网安备 33010602011771号