易基因:癌症中的cfDNA羟甲基化:检测方法及临床应用 | 精准医学
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
在精准肿瘤学时代,异常的遗传和表观遗传学变化鉴定促进了癌症诊断、管理和治疗方式的改变。DNA羟甲基化(5-羟甲基胞嘧啶,5hmC)是一种新兴的表观遗传修饰,由TET酶通过氧化5-甲基胞嘧啶(5mC)形成。DNA羟甲基化表现出组织和癌症特异性模式,在DNA去甲基化和基因调控中至关重要。5hmC检测方法的最新进展,以及在游离细胞DNA(Cell-Free DNA,cfDNA)中鉴定5hmC,表明了游离细胞5hmC作为癌症生物标志物的潜力。本文探讨了DNA(尤其是cfDNA)羟甲基化在癌症中的当前和新兴技术及应用。
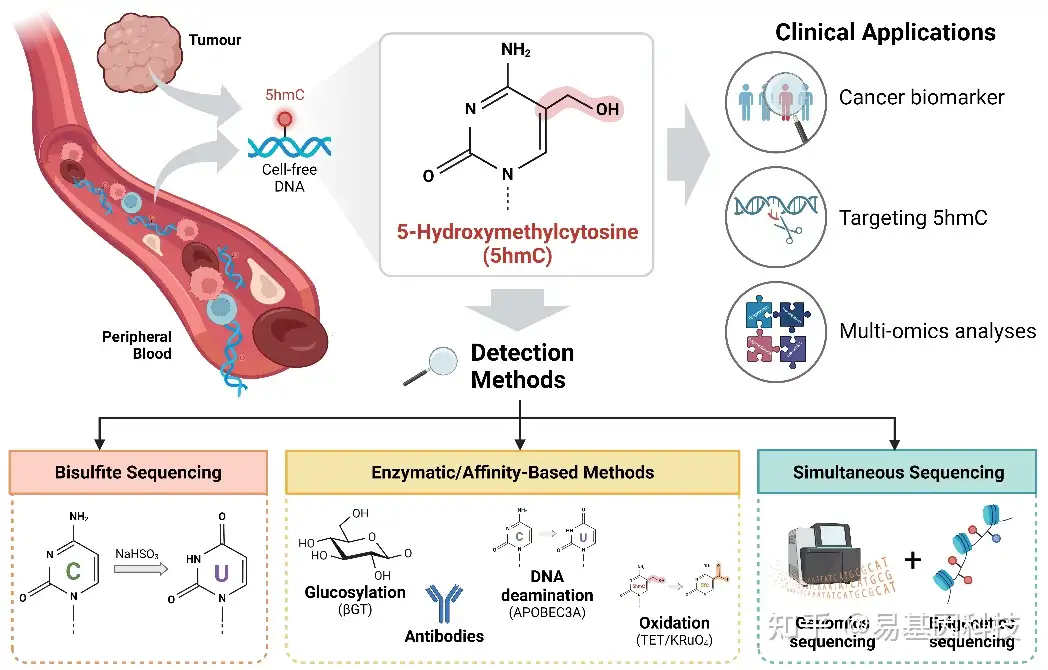
5hmC是一种在正常发育和基因调控等生物过程中发挥着重要作用的表观遗传修饰。5hmC失调会介导各种疾病(如癌症)发展,这表明了其生物学过程中重要作用。液体活检为获得基因组和表观基因组数据提供了无创方法,彻底改变了癌症的筛查和治疗方式。通过研究来自血浆等体液中的cfDNA,特别是循环肿瘤DNA(circulating tumour DNA,ctDNA)为捕捉肿瘤的异质性、监测遗传和表观遗传动态以及检测癌症或复发的早期迹象提供了机会。以下内容从DNA羟甲基化的生物学角色和机制进行概述,并为DNA羟甲基化作为癌症生物标志物的最新进展介绍了更新视角。最后讨论了在5hmC研究中使用cfDNA和综合多组学分析,并讨论其在临床应用中的机遇和挑战。
DNA羟甲基化的生物学功能和机制
DNA去甲基化通路中的羟甲基化
DNA甲基化(5-甲基胞嘧啶,5mC)是哺乳动物DNA中研究最为深入的表观遗传标记,对基因调控至关重要。5mC可以通过DNA甲基转移酶(DNMT)从S-腺苷甲硫氨酸(SAM)向胞嘧啶的5位共价转移甲基基团来酶促产生。在主动去甲基化通路中,DNA 5hmC通过TET双加氧酶(例如TET1、TET2和TET3)氧化5mC产生(图1)。这一反应需要α-酮戊二酸(α-KG)和氧气,在铁和抗坏血酸(维生素C)下产生5hmC、琥珀酸和二氧化碳。TET酶进一步氧化5hmC产生5-甲酰胞嘧啶(5fC)和5-羧基胞嘧啶(5caC),尽管这些中间体的存在于5hmC的频率要低得多。与5hmC不同,5fC和5caC不稳定,分别被胸腺嘧啶DNA糖基化酶(TDG)和基础切除修复(BER)机制切除和修复,恢复为未修饰的胞嘧啶形式。5hmC也可以通过激活诱导的胞嘧啶脱氨酶(AID)/载脂蛋白B mRNA编辑酶,催化多肽类(APOBEC)脱氨成5-羟甲基尿嘧啶(5mU),然后通过BER去甲基化。通过复制稀释也可以发生胞嘧啶修饰的被动去甲基化。
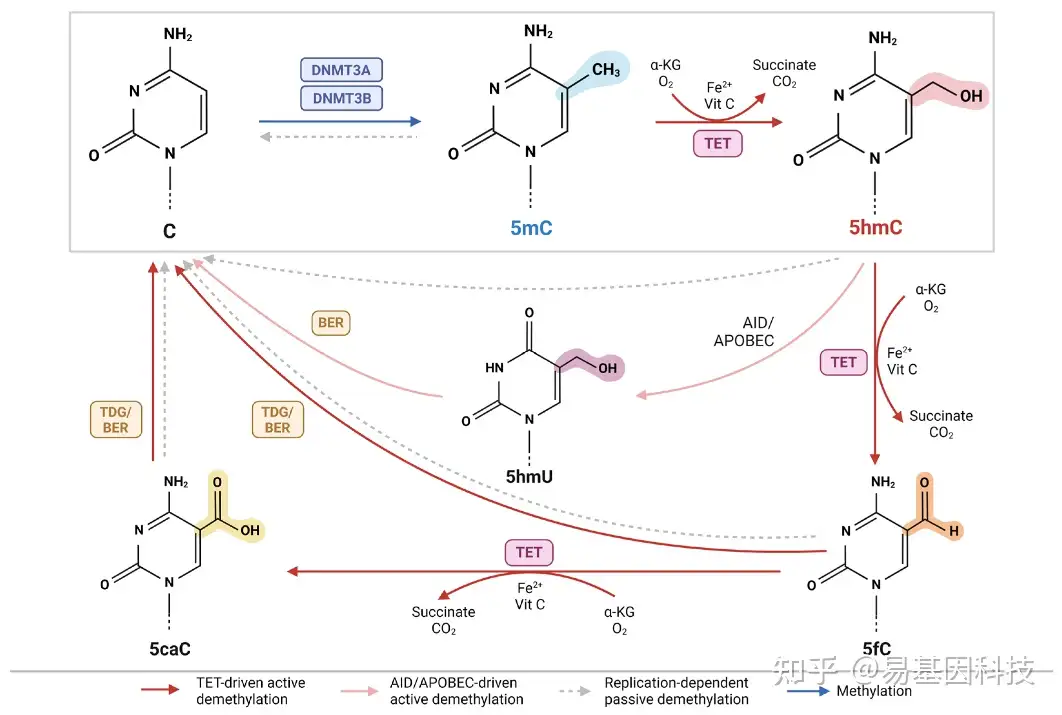
图1:活性DNA去甲基化通路。
与5mC、5fC和5caC相比,TET酶和TDG-BER耦联机制对5hmC的亲和力较低,表明5hmC是稳定的,并不容易去甲基化。此外,5hmC可以在多次细胞分裂中持续存在,表明它可能在去甲基化通路之外具有重要的生物学功能。
DNA 羟甲基化的生物学功能和分布
5hmC在胚胎发育、干细胞多能性、T淋巴细胞(T细胞)的缺失和分化、以及不同疾病的病因学等多种生物过程中发挥重要作用,并在活跃的基因调控中扮演着关键角色。5hmC主要在转录活跃区域中富集,如基因体、增强子以及启动子周围(图2)。富含5hmC区域通常位于活跃的组蛋白修饰(例如H3K4me2和H3K4me3)和组蛋白标记的增强子(例如H3K4me1和H3K27ac)附近,表明它可能参与调节活跃的基因表达(图2)。许多研究表明5hmC水平与基因表达之间存在正相关。
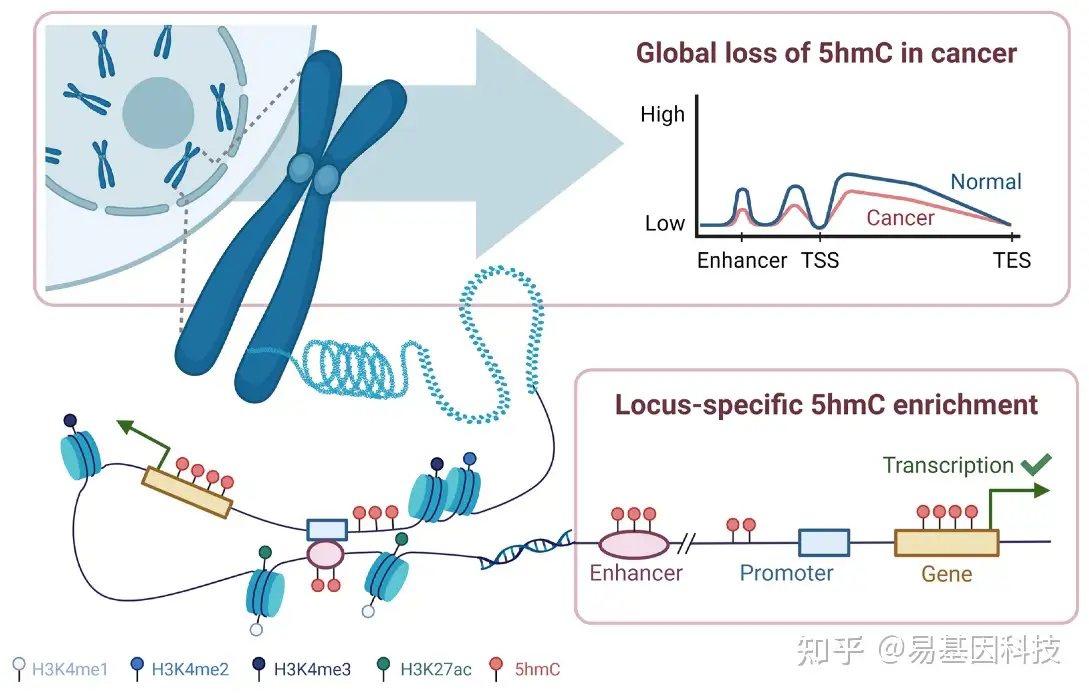
图2:全基因组和位点特异性DNA羟甲基化模式。
尽管全基因组5hmC水平较低,但其在不同细胞类型、组织和疾病状态下的分布差异很大。一般来说,5hmC在脑组织(约占所有胞嘧啶的0.67%)和胚胎干细胞中最普遍。5hmC丰度似乎与细胞增殖呈负相关,后有丝分裂细胞通常具有较高的5hmC水平。由于这些细胞往往具有高度的增殖性,导致在细胞系和肿瘤组织中检测到的5hmC水平较低。
游离DNA和羟甲基化组
对游离细胞羟甲基化的研究开启于2017年,song等人开发了一种高度敏感的化学标记检测方法,能够检测到少量cfDNA输入中的低水平5hmC。从那时起开始在各种临床应用中被研究,包括早期癌症检测和预测癌症结果。尽管与组织相比,cfDNA中的5hmC水平通常较低,但先前的研究表明cfDNA 5hmC模式和动态反映了组织中发生变化。此外,由于5hmC相对稳定,基因组DNA上的5hmC标记可以在cfDNA片段中重现,表明它有潜力作为一种微创的癌症生物标志物。
癌症中的DNA羟甲基化模式
DNA羟甲基化模式可以在全局(全基因组)或局部(位点特异性基因组,图2)范围内进行评估。在各种类型的癌症中,包括膀胱、脑部、胰腺和乳腺癌,都检测到全基因组5hmC丢失。在某些情况下,5hmC水平更大降低与更侵袭性肿瘤和更差预后相关。在皮肤T细胞淋巴瘤(CTCL)细胞系中,Qiu等人(2018年)观察到随着肿瘤侵袭性增加,5hmC逐渐丢失,其中大细胞转化的CTCL细胞系显示出最大的5hmC百分比降低,其次是未转化的CTCL和前CTCL斑块和补丁。同样,Song等人(2017年)发现在非小细胞肺癌(NSCLC)中,cfDNA中的5hmC呈现阶段依赖性缺失,其中转移性NSCLC显示出与非转移性NSCLC和健康个体相比,归一化羟甲基化区域的最低水平。但这些发现可能因所研究的组织类型和人群而异。例如,与Song等人(2017年)的研究不同,其他在中国人群中进行的NSCLC研究报告称,与健康对照组相比,癌症中的5hmC水平全基因组增加。尽管如此,整体5hmC模式在癌症和非癌症对照之间显示出不同的特征。
癌症中也在转录活跃区域描述了特定基因位点的5hmC增加。在结直肠癌的cfDNA研究中,Li等人(2017年)报告了5hmC在基因体、开放染色质区域(DNaseI高敏感位点)以及带有活跃组蛋白修饰(H3K27ac,H3K4me1,H3K9me1)的区域富集。在另一项研究中,Sjöstrom等人(2022年)揭示了前列腺癌驱动基因的增强子或转录起始位点上游的局部5hmC富集,尤其是雄激素受体(AR)和叉头盒蛋白A1(FOXA1),与其基因表达相关。表明5hmC失调可能导致疾病状态,即使在驱动基因上没有遗传突变。
总的来说,这些研究表明整体和局部5hmC模式是强大的标记物,能够区分癌症和非癌症样本,暗示了5hmC作为癌症生物标志物的潜力,尽管还需要进一步研究。
羟甲基化检测方法的演变
在过去十年中,对羟甲基化组的定量和分析取得了显著进展。早期5hmC分析研究主要通过质谱、薄层色谱、高效液相色谱(HPLC)、免疫检测和光谱技术等定量方法。虽然这些方法能够准确定量5hmC水平,但其样本制备复杂性、高昂的设备和试剂成本以及低分辨率使得它们在临床环境中难以实施。自那以后,科学家们开发出了基于测序的方法,用于全基因组和单碱基分辨率的分析。表1总结了最常见的5hmC分析技术。
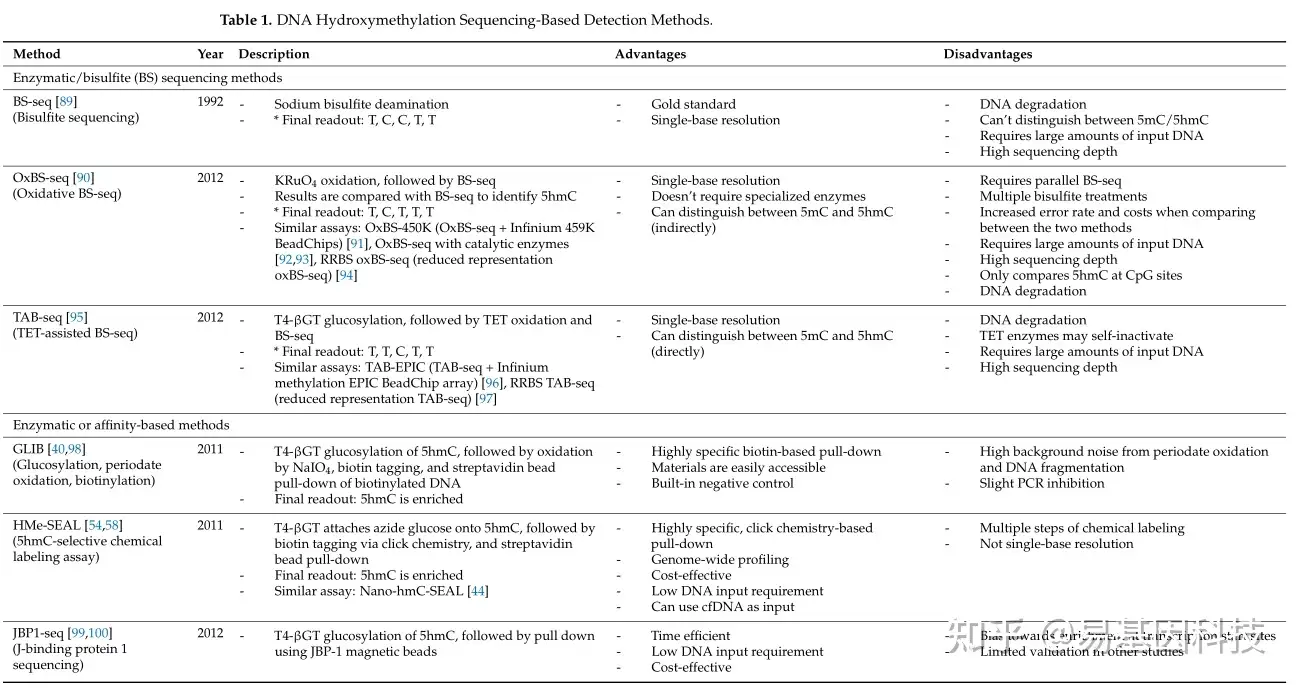

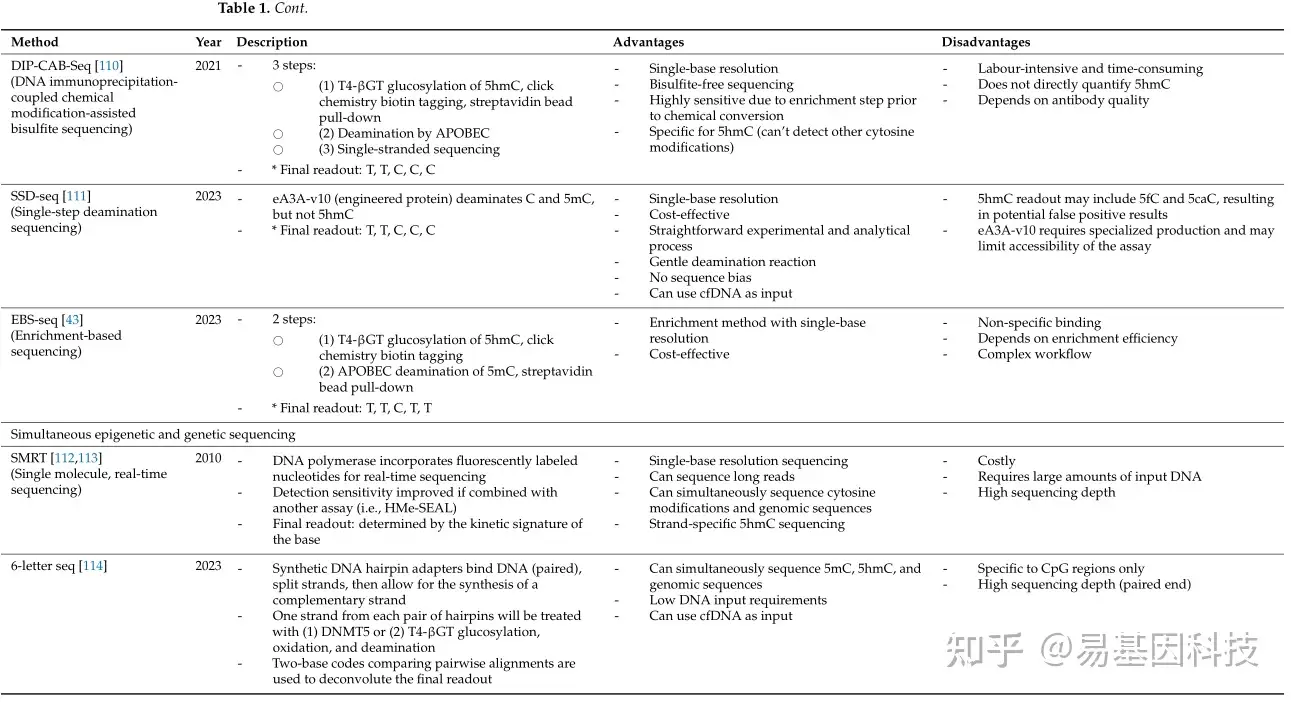
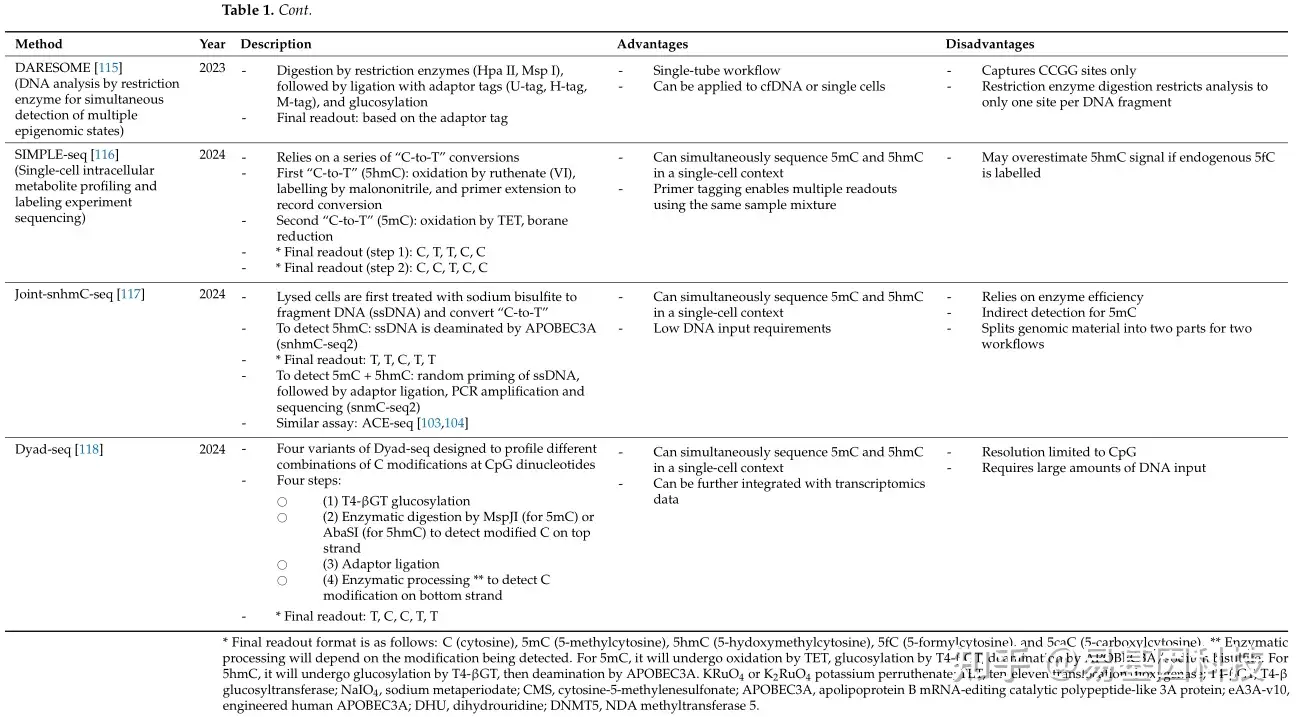
表1:基于DNA羟甲基化测序的检测方法。
亚硫酸盐测序方法
亚硫酸盐测序(BS-seq)是DNA甲基化分析的黄金标准方法,能够提供全基因组范围内的单碱基分辨率评估。传统的BS-seq用亚硫酸钠处理DNA,将未修饰的胞嘧啶(C)、5fC和5caC脱氨基成尿嘧啶(U),并将5hmC转化为胞嘧啶-5-甲磺酸(CMS),而5mC保持不变。由于5mC和CMS的最终读数(readout)都是胞嘧啶,BS-seq的一个限制是无法区分5mC和5hmC。为了克服这一障碍,可以在亚硫酸盐处理前添加化学物质来保护或修饰碱基,从而实现对甲基化状态的更精确区分。 氧化亚硫酸盐测序(OxBS-seq)和TET辅助亚硫酸盐测序(TAB-seq)是改良亚硫酸盐技术的示例。OxBS-seq利用高碘酸钾(KRuO4)将5hmC氧化成5fC。由于5fC是一种不稳定的中间体,TDG和BER机制会将其再转化为胞嘧啶,使其在亚硫酸盐处理下发生脱氨基。与传统亚硫酸盐测序读数比较,可以识别5hmC(BS-seq中的“C”和OxBS-seq中的“T”)。TAB-seq依赖于T4噬菌体β-葡萄糖基转移酶(T4-βGT)将葡萄糖基团转移到5hmC上,保护其不受TET1氧化。经过氧化和亚硫酸盐处理后,最终读数(readout)允许直接识别5hmC(5hmC读作“C”,而胞嘧啶、5mC和其他氧化衍生物读作“T”)。
酶促和亲和方法
为了克服基于亚硫酸盐方法的局限性,非亚硫酸盐技术使用化学物质、酶和抗体来葡萄糖基化、脱氨基、氧化或选择性地拉下5hmC,以捕获其全基因组分布。这些策略反过来提高了5hmC分析技术的灵敏度、特异性和可行性,为临床应用的过渡提供了便利。
- 化学捕获和葡萄糖基化技术
葡萄糖基化涉及通过T4-βGT将葡萄糖基团转移到5hmC的羟基上,形成葡萄糖基-5hmC(5ghmC)。葡萄糖分子可以被放射性标记以进行定量,用于保护5hmC免受氧化,或者用于将各种标签附着在5hmC上。例如,葡萄糖基化、周期酸氧化、生物素化(GLIB)使用葡萄糖基化来保护5hmC,然后再通过NaIO4氧化。随后进行生物素化和链霉亲和素珠子捕获。GLIB有几个局限性,包括由NaIO4氧化和DNA片段化引起的高背景噪声,以及对PCR的轻微抑制作用。这些可能影响后续分析或在测序过程中低估富含5hmC区域。尽管这些局限性可以通过实验的预期阴性对照(无T4-βGT的条件)和用Helicose仪器替换PCR来解决,但这些解决方案可能会产生额外成本,并且可能不是所有实验室都能轻易获得。
5hmC选择性化学标记(HMe-SEAL)是另一种基于葡萄糖基化的技术,它利用T4-βGT将修饰的叠氮葡萄糖分子转移到5hmC上。随后通过点击化学添加生物素标签,以及随后的链霉亲和素珠子拉下生物素化片段。目前,HMe-SEAL优化版本广泛用于稀有细胞群体(Nano-hmC-Seal)和游离细胞DNA的5hmC分析。然而,像其他富集技术一样,HMe-SEAL无法对5hmC位点进行单碱基分辨率分析,限制其定量5hmC水平。尽管如此,HMe-SEAL为cfDNA研究提供了一个机会,因为它需要的DNA起始量低,并且可以有效地富集含有5hmC的片段,从而实现更具成本效益的测序。
同样,J结合蛋白1测序(JBP1-seq)也依赖于T4-βGT来葡萄糖基化5hmC。J结合蛋白1是葡萄糖基化5hmC的天然抗体。在一项比较JBP1-seq与HMe-SEAL和其他基于亲和性的5hmC分析方法的研究中,JBP1-seq显示出显著不同的5hmC谱图,并且无法将染色质状态与5hmC模式相关联。此外,它可能偏向于转录起始位点,导致5hmC的富集度降低。然而,优化的JBP1-seq工作流程已经展示了改进的5hmC富集能力,更快的性能(4.5小时)和降低的DNA起始量要求。需要进一步验证以全面评估这种优化工作流程的性能。
其他葡萄糖基化方法包括Jump-seq和5hmC连接寡核苷酸引导测序(hmTOP-seq)。Jump-seq和hmTOP-seq都使用T4-βGT将叠氮葡萄糖附着在5hmC上,然后通过点击化学标记带有发夹或连接的寡核苷酸,并延伸引物以定位5hmC。最近,hmTOP-seq也被用于检测cfDNA中的5hmC。尽管研究围绕母血cfDNA的产前检测展开,hmTOP-seq在极低测序深度下检测胎儿唐氏综合症的准确性达到了100%。表明它有潜力成为分析游离细胞羟甲基化组的工具。然而,需要注意的是,Jump-seq和hmTOP-seq都依赖于间接方法来定量5hmC,可能会在测序后引入偏差和假象。此外,Jump-seq往往对高CpG密度区域有偏倚,如果5hmC峰太接近可能会表现不佳。
②DNA脱氨方法
AID/APOBEC酶是一种替代的DNA脱氨方法,无需亚硫酸盐处理过程中产生的不稳定的磺化中间体。这一反应使用锌辅因子来脱氨基单链DNA上的胞嘧啶。第一个采用这种策略的技术是APOBEC偶联表观遗传测序(ACE-seq)。在ACE-seq中,5hmC首先通过T4-βGT葡萄糖基化以保护其不受随后APOBEC3A脱氨基的影响。另一方面,5mC和其他未修饰的胞嘧啶将在APOBEC3A处理下被脱氨基。ACE-seq的一个主要优势是,ACE-seq可以实现5hmC的单碱基分辨率比对,所需的DNA 起始量比BS-seq少1000倍。酶促甲基测序(EM-seq)遵循与ACE-seq相同的工作流程,但在葡萄糖基化之前使用TET2氧化5mC和5hmC。
单步脱氨基测序(SSD-seq)是另一种脱氨基技术,它利用特别设计的蛋白eA3A-v10来脱氨基胞嘧啶和5mC,但不脱氨基5hmC。这种方法消除了脱氨基前额外的葡萄糖基化或氧化步骤的需要。此外,eA3A-v10的脱氨基使用温和的脱氨基反应,克服了亚硫酸盐处理对DNA损伤的限制。像ACE-seq一样,需要注意的是5hmC的reads可能包括5fC和5caC的痕迹,这可能会高估5hmC水平。然而,由于它们的普遍性和稳定性远低于5hmC,可能不会对最终结果产生显著影响。
新的脱氨基方法将脱氨基与现有的分析技术相结合,以提高全基因组5hmC检测的分辨率。DNA免疫沉淀偶联化学修饰辅助亚硫酸盐测序(DIP-CAB-seq)是一种三步策略:(1)葡萄糖基化、点击化学和生物素-链霉亲和素互作,如HMe-SEAL,(2)APOBEC脱氨基,以及(3)单链测序。尽管DIP-CAB-Seq工作量大,但它展示了与HMe-SEAL相当的5hmC富集能力,同时实现了改进的单碱基分辨率检测。同样,基于富集的单碱基分辨率5hmC测序(EBS-seq)遵循类似的工作流程,其中APOBEC脱氨基发生在链霉亲和素珠子拉下之前。即使在5hmC含量低的样本中,如人类癌细胞系和组织,也能有效地检测到5hmC。- 基于氧化策略
基于氧化的方法,不使用亚硫酸盐,通常用于在化学处理之前转换或保护胞嘧啶修饰。例如,化学辅助的5hmC到T的转换测序(hmC-CATCH)使用高碘酸钾(KRuO4)将5hmC氧化并转化为5fC,然后再用indandione进行标记。这个过程确保了只有标记了5fC的产物在PCR过程中发生“C到T”的转变,而其他胞嘧啶衍生物保持不变。另一种技术,TET辅助的吡啶硼烷测序(TAPS-seq),使用TET将5mC和5hmC氧化成5caC。与TAB-seq不同,它使用吡啶硼烷将5caC还原为二氢尿嘧啶(DHU),而不是亚硫酸盐处理。当与TAPSβ-seq平行进行时,后者结合了葡萄糖基化以保护5hmC免受氧化,它们的集体reads允许识别5hmC。这种检测需要在酸性条件下长时间孵育,但它比亚硫酸盐处理的破坏性小。
④ 基于抗体方法
特异性靶向5hmC或其衍生物的抗体也可以用来捕获羟甲基化DNA片段。羟甲基化DNA免疫沉淀(hmeDIP)利用单克隆抗5hmC抗体来捕获和拉下羟甲基化DNA,已广泛用于癌症组织和细胞系中5hmC的局部和全基因组分析。尽管基于抗体的分析技术受到抗体结合效率的变异性限制,可能偏向于高羟甲基化区域,但它们具有成本效益、易于实施,并且可以对5hmC具有高度特异性。
DNA羟甲基化及其临床应用
5hmC作为一种有前途的癌症生物标志物,展现出独特的全局和局部模式,能够区分癌症和非癌症样本。以往的研究已经探索了它作为癌症的诊断、预后和预测生物标志物的潜力,强调了它在癌症管理中的实用性。图3和表2突出了5hmC作为癌症生物标志物的临床应用示例。
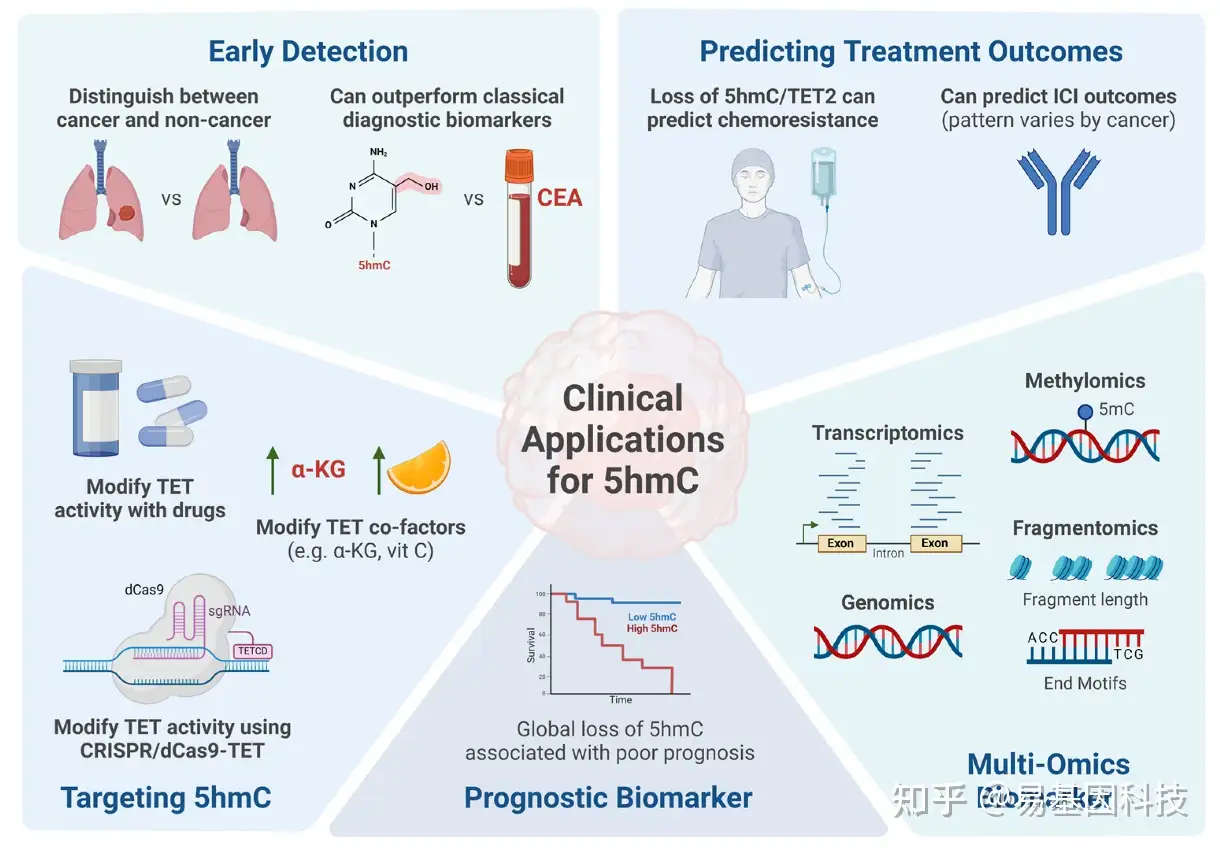
图3:DNA羟甲基化在癌症中的临床应用。

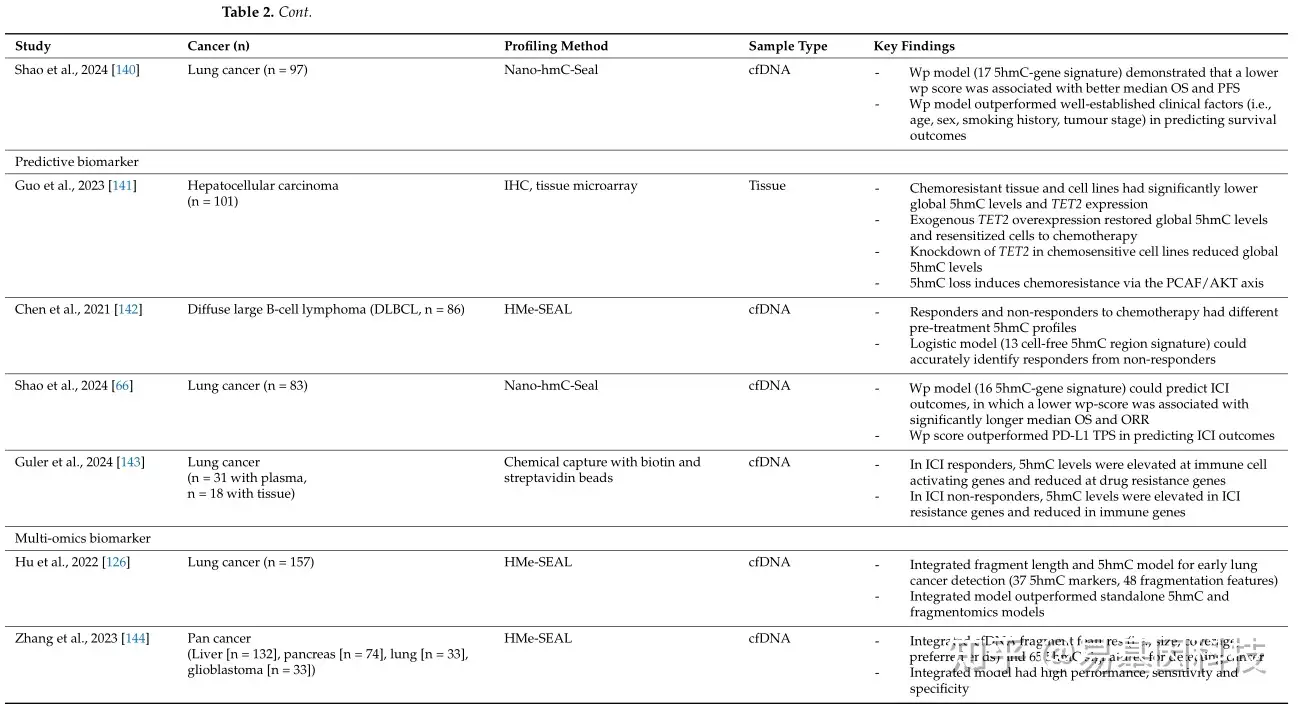

表2:DNA羟甲基化作为癌症生物标志物示例。
DNA羟甲基化作为癌症检测的生物标志物
早期发现癌症可以使得治疗更加有效,预后更好,因此拥有可靠的诊断生物标志物至关重要。然而,由于早期肿瘤含量极低,尤其是循环肿瘤DNA(ctDNA),这使得检测变得具有挑战性。整体和特定基因位点的5hmC模式是很有前景的诊断生物标志物。
在一项泛癌研究中,Shao等人(2022年)利用健康个体和患有膀胱、乳腺、结直肠、肾脏、肺或前列腺癌患者的cfDNA建立了一个24个5hmC基因模型。他们的模型在验证队列中成功地区分了癌症和健康个体(AUC=91%,灵敏度=68.6%,特异性=96.6%)。当单独考虑每种癌症时,他们的癌症特异性加权诊断(wd)模型实现了从94%到99.8%的AUC,灵敏度从80%到96%,特异性从96%到100%。对于早期疾病的检测性能也很高,泛癌灵敏度在71.4%和81.3%之间,癌症特异性标志物在I期和II期疾病中分别为89.3%和94.1%。
在一项多中心研究METHOD-2(NCT03676075)中,Chang等人(2024年)还开发了一个能够区分I-III期结直肠癌患者和非癌症对照的cfDNA 5hmC分类器。他们基于96个5hmC基因体的模型在内部验证中实现了94.3%的AUC,在外部验证中实现了90.7%的AUC。他们的cfDNA 5hmC基础分类器的性能超过了经典的结直肠癌生物标志物癌胚抗原(CEA),后者在内部和外部验证队列中的AUC分别为77.1%和73.2%。在90%的特异性下,基于5hmC的评分在I期和II/III期结直肠癌中的灵敏度为73.5%和85.3%,而CEA在同一阶段的灵敏度分别为29.4%和47.2%。在肝细胞癌、肺癌和结直肠癌的研究也表明,基于5hmC的模型可以超越传统的诊断生物标志物,突显了5hmC在癌症检测中的潜力。
尽管全局和基因特异性的cfDNA 5hmC模式作为癌症的诊断生物标志物具有潜力,甚至是对于早期疾病,但在这些模型可以转化为临床实践之前,仍有几个问题需要解决。首先,当前的诊断模型往往只关注基因特异性的5hmC标记,而其他富含5hmC的调控因子,如增强子,往往被忽视。将其他调控因子纳入这些模型可以提高性能并改善我们对观察到的5hmC模式背后的生物学的理解。其次,未来的研究应该优先验证现有的基因标记模型,特别是在不同的地理和种族群体中,以评估这些模型的普适性。
DNA羟甲基化在癌症中的预后价值
除了早期检测,有证据支持5hmC分析在癌症预后中的价值。例如,在肝内胆管癌(ICC)中,Dong等人(2015年)表明,与高/阳性5hmC水平相比,组织中低或阴性5hmC水平与更高的肿瘤阶段、更多的淋巴结转移、更短的总生存期和更差的无病生存期相关。同样,Kuang等人(2024年)揭示了TET2/5hmC子宫内膜腺癌与其他TET2和5hmC表达组合相比,与最差的总生存期相关(p<0.001)。相比之下,TET2/5hmC子宫内膜癌与分化良好的细胞、最小的肌层侵袭、阴性淋巴结转移和较低的肿瘤阶段显著相关。多变量分析进一步表明,TET2/5hmC关系可以作为子宫内膜癌的独立预后因子(HR:2.84,95%CI 1.23–3.61,p=0.007)。同样,在胃癌中,Fu等人(2022年)证明高5hmC是OS(风险比[HR]=0.61,95%CI0.38–0.98,p=0.04)和较低肿瘤阶段(HR=0.32,95%CI0.13–0.77,p=0.011)的独立、有利预测因子。
游离细胞5hmC标记也可用于癌症预后或患者风险分层。在弥漫性大B细胞淋巴瘤(DLBCL)中,Chiu等人(2019年)创建了一个基于29个5hmC基因的加权预后(wp)模型,该模型在预测无进展生存期(EFS)和OS方面实现了96%的准确性、86%的灵敏度和100%的特异性。高加权预后(wp-score)的患者与低wp-score的患者相比,OS和EFS显著更差。5hmC模型的灵敏度、特异性和准确性超过了现有的DLBCL预后因子,如乳酸脱氢酶水平升高、激活的B细胞型DLCBL和高国际预后指数(准确性=36–81%,灵敏度=56–80%,特异性=29–89%)。
在肝细胞癌(HCC)中,Cai等人(2021年)使用cfDNA 5hmC标记和两个HCC蛋白生物标志物(甲胎蛋白[AFP]和去γ-羧基凝血酶原[DCP],表2)开发了一个HCC评分。高HCC评分与HCC的临床病理预后因子,如微血管侵犯(MVI)、肿瘤大小和肿瘤阶段,呈正相关,并且与更高的复发率、更短的无复发生存期和更短的OS显著相关。同时,HCC评分也与实时肿瘤负担动态正相关,表明其预测疾病复发的潜力。然而,这一发现受到研究样本量小的限制。
在另一项研究中,Shao等人(2023年)使用基于H3K4me3标记的前500个最可变cfDNA 5hmC区域的无监督分层聚类,将54名急性髓系白血病(AML)患者分为三个聚类。第三聚类在TET2启动子中表现出最低的5hmC水平,最短的OS和最高的白血病负担。第三聚类和第一聚类之间的差异性羟甲基化区域(DhMR)富集在与不良预后相关的细胞生存/增殖通路中(例如,mTOR、ERK/MAPK和胰岛素受体信号通路)。
同样,在非小细胞肺癌(NSCLC)中,Shao等人(2024年)使用17个5hmC基因标记构建一个预后模型。低wp-score患者与高wp-score患者相比,平均OS(18.8vs.5.2个月,p=0.0006;HR0.22,95%CI0.09–0.57)和平均PFS(8.8vs.3.3个月,p=0.054;HR0.45;95%CI0.20–1.00)显著更长。预后评分在预测生存结果方面比已知的临床因子(如年龄、性别、吸烟史和肿瘤阶段)更准确。
总的来说,这些研究表明游离细胞和基于组织的5hmC模式可以作为癌症的预后生物标志物。对这些5hmC模式背后的基因和通路进行进一步的研究,将为导致不良预后风险因子提供更深入的见解。
DNA羟甲基化整合的多组学分析
癌症是几个潜在生物网络的总和,而不仅仅是个体现象。因此,研究DNA 5hmC与其他生物学层面(如甲基化组学、片段组学和转录组学)的整合和互作日益增加。特别是研究cfDNA片段特征(片段组学),如大小、末端基序、锯齿末端、核小体定位和覆盖范围,可以揭示肿瘤和非肿瘤DNA之间的差异。例如,ctDNA片段长度通常比cfDNA片段短(ctDNA中约145bp,cfDNA中约166bp)。在非小细胞肺癌(NSCLC)中,Hu等人(2022年)开发了一个整合了5hmC和片段组学模型的早期检测模型。他们使用37个cfDNA 5hmC标记和48个片段特征(在不同窗口中短片段与长片段的比例),该模型能够区分NSCLC患者和非癌症对照组,并在两个验证队列中实现了86-94%的AUC值,83-88%的灵敏度和78-90%的特异性。此外,这个整合模型的表现优于单独的5hmC和片段模型,突出了将片段组学长度与cfDNA 5hmC结合用于癌症检测的价值。在一项泛癌研究中,Zhang等人(2023年)使用5hmC标记和多个cfDNA片段特征(包括大小、覆盖范围和优选末端)建立了一个诊断模型。该模型实现了高AUC值、灵敏度和特异性,展示了整合5hmC和片段特征用于癌症诊断的潜力。整合的5hmC和片段组学模型也已在其他癌症的早期检测中进行了研究。
以前的研究小组还探索了5mC、5hmC和基因表达之间的互作,以更深入地理解癌症背后的遗传和表观遗传模式。Shi等人(2023年)结合了基因组、转录组、甲基化组和羟甲基化组的数据,以更好地理解膀胱癌复发的发展。他们证明尽管膀胱癌没有已知的驱动突变,但5hmC或5mC诱导的转录变化与膀胱癌复发和肿瘤免疫逃逸相关通路有关。表明表观遗传改变在膀胱癌复发中可能比基因组突变发挥更显著的作用。在儿童中枢神经系统(CNS)癌症中,Lee等人(2024年)整合了5mC、5hmC和单核RNA-seq数据,以研究细胞类型组成对表观遗传学的影响。作者发现5mC和5hmC的丰度和变化受到肿瘤组成和异质性的强烈影响。他们还观察到在肿瘤组织中5mC和5hmC的全基因组失调的普遍性,尽管5hmC在基因组中的丰度明显低于5mC,这表明5hmC在肿瘤发生中具有独立且至关重要的作用。
因此,将5hmC与其他组学数据集整合为增强当前癌症生物标志物提供了机会,并加深了对5hmC模式和癌症病理生理学的理解。
DNA羟甲基化靶点作为癌症潜在治疗策略
尽管已有多项研究报道了癌症中5hmC的全局和局部模式,但推动这些变化的具体机制仍然难以捉摸。5hmC水平的降低与TET活性受损有关,这可能是由于TET基因的失活突变、TET表达下调以及TET辅因子和底物抑制。先前的研究表明,过表达TET可以显著抑制细胞增殖、迁移和侵袭,而敲低TET则表现出相反的表型。因此,靶向TET可能为癌症治疗提供一种潜在策略。目前还没有批准的药物专门靶向TET蛋白进行癌症治疗。然而,先前的急性髓系白血病(AML)药物筛选研究揭示了如NSC-311068和NSC-370284等化合物,它们可以抑制TET1表达,进而降低5hmC水平,以抑制体外和体内的细胞活力和肿瘤进展。另一种新兴策略涉及使用CRISPR/cas9表观基因组编辑工具,其中去活化的cas9(dcas9)与TET蛋白(TETCD)的催化域融合,以靶向并去甲基化由引导RNA(sgRNA)指定基因区域的DNA。Choudhury等人(2016年)使用CRISPR/dCas9-TET1CD系统进行的初步工作展示了体外BRCA1启动子的成功去甲基化,从而恢复BRCA1表达,并抑制了多种癌细胞系的细胞生长。同样,Xu等人(2018年)使用与TET3CD耦合的高保真度dcas9去甲基化肾脏纤维化中的高甲基化抗纤维化基因,展示了体外和体内抗纤维化基因Rasal1和Klotho的成功基因特异性重新激活。未来的研究方向,如优化sgRNA结合位点相对于目标的位置和研究脱靶效应,将提高这种CRISPR/Cas9技术的广度和实用性。TET也可以通过改变其底物和辅因子的可用性间接修饰。例如,α-酮戊二酸(α-KG)是三羧酸循环(TCA)中的关键中间体,也是TET在去甲基化途径中的底物。Liu等人(2023年)的研究表明,α-KG补充增加了黑色素瘤中TET2/3的活性,导致PD-L1启动子处5hmC水平升高,提高了ICI的疗效。异柠檬酸脱氢酶(IDH)基因IDH1和IDH2的突变也可能影响TET活性,因为突变酶将α-KG转化为2-羟基戊二酸,这是TET催化位点的竞争性抑制剂。然而,IDH突变是否是癌症中5hmC耗竭的主要原因仍不确定。维生素C是另一种可以增强TET活性的辅因子,从而增加5hmC水平。Peng等人(2018年)证明,维生素C处理可以以时间和浓度依赖性的方式恢复5hmC水平,同时抑制膀胱和肾脏癌细胞系的细胞增殖并诱导凋亡。尽管TET活性增加,但TET1/2/3的表达水平并没有显著变化,表明维生素C增强了TET活性,而不是表达水平。在其他癌症中也观察到了类似的发现,表明维生素C治疗可能是癌症的潜在选择。 尽管α-KG和维生素C在改变TET活性方面显示出潜力,但这些辅因子也参与多种细胞过程,因此可能导致意想不到的副作用。需要进一步研究这些辅因子改变的确切机制和后果。
挑战和未来方向
选择正确的5hmC检测方法
5hmC检测方法进步导致了研究羟甲基化组的全基因组分析策略的改进。目前的技术现在可以区分各种胞嘧啶修饰(表1),从而可以单独评估每种修饰在肿瘤发生中的作用。同时对基因组和表观基因组进行测序的新方法具有巨大的前景,为使用单一工作流程进行多组学分析提供了机会。然而,DNA输入要求和测序成本等挑战依然存在。此外,随着cfDNA输入的使用不断增长,即使在低浓度下,也必须有能够检测5hmC的灵敏检测方法。尽管如此,基于液体活检技术的发展正在出现,可以提高cfDNA检测的灵敏度和特异性。方法的选择可能会受到分析解决这些障碍的程度或所选方法所需试剂的可用性的影响。
cfDNA羟甲基化的多组学分析
对癌症检测、预后和预测治疗结果的cfDNA羟甲基化组进行分析正在取得进展。将5hmC数据与其他组学相结合,不仅可以补充当前的生物标志物模型,还可以更全面地了解癌症生物学。然而,在临床实施可行之前,需要解决几个挑战。
首先,每个血浆样本中ctDNA含量有限(5-10ng/mL),这对多组学研究构成挑战,因为进行多次检测时ctDNA含量往往不足。
其次,多组学cfDNA研究的标准化方法仍然缺乏,因为这仍然是一个不断发展的领域。开源生物信息学管道、ctDNA解卷积方法和合成参考对照(即DNA尖峰)的开发将有助于确保集成数据的可重复性和可比性。
第三,来自肿瘤微环境或外周血白细胞中cfDNA的表观遗传信号可能会破坏或稀释正在研究的ctDNA信号。因此,至关重要的是要有适当的工具来细化肿瘤特异性5hmC信号,同时滤除背景、非肿瘤噪声。目前的工具包括calling ctDNA变体和过滤外周血白细胞信号。
最后,拥有适当的生物信息学工具和基础设施对于支持新发现的整合和分析至关重要。
结论
DNA羟甲基化作为癌症检测、预后和治疗反应预测的生物标志物具有巨大的前景。分析方法的进展,特别是cfDNA方法的进步,使人们能够开发出高灵敏度和特异性的工具来检测羟甲基化。通过将5hmC数据与其他多组学数据集整合,可以进一步增强这些发展,提供对癌症机制的更全面了解,并有助于发现新的生物标志物和治疗靶点。然而,在更广泛地临床采用5hmC生物标志物之前,仍有几个挑战需要解决。这些包括进一步验证当前的生物标志物,解决测序和输入相关的问题,以及建立必要的生物信息学工具和分析基础设施。尽管如此,随着对5hmC机制和检测方法的持续研究,5hmC有可能成为改善癌症治疗的有价值的生物标志物。
易基因提供全面的表观基因组学(DNA甲基化、DNA羟甲基化)和表观转录组学(m6A、m5C、m1A、m7G、ac4C)、染色质结构与功能组学技术方案(ChIP-seq、ATAC-seq),详询易基因:0755-28317900。

参考文献:
Li JJN, Liu G, Lok BH. Cell-Free DNA Hydroxymethylation in Cancer: Current and Emerging Detection Methods and Clinical Applications. Genes (Basel). 2024 Sep 3;15(9) pii: genes15091160. doi: 10.3390/genes15091160. PubMed PMID: 39336751.
相关阅读:
深度综述 | cfDNA甲基化诊断和监测肿瘤的研究进展与展望:胰腺癌

 浙公网安备 33010602011771号
浙公网安备 33010602011771号