易基因|糖尿病大血管和微血管病变的表观遗传学调控机制(DNA甲基化+组蛋白修饰):综述
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
2022年12月09日,《Frontiers in Endocrinology》杂志发表了题为“Epigenetic basis of diabetic vasculopathy”的综述文章,综述了表观遗传学修饰如DNA甲基化和组蛋白修饰在2型糖尿病(Type 2 diabetes mellitus,T2DM)大血管和微血管并发症病变中的重要作用。

期刊:Frontiers in Endocrinology / Front Endocrinol (Lausanne)
日期:2022.12
IF:6.055 /Q2
摘要
2型糖尿病(T2DM)会引发外周血管疾病,其中包括重要营养素在内多种血液传播因子无法到达受影响的组织。组织表观基因组对慢性高血糖症敏感,可引起微血管和大血管并发症的发病机制。T2DM的血管并发症可能会导致器官功能失调,而糖尿病的主要负担就在于各种并发症,其中一大重要并发症就是难以愈合的糖尿病溃疡。因此当前的研究必须有助于识别更有效诊断和治疗早期血管损伤的方法。本文综述了表观遗传过程如DNA甲基化和组蛋白修饰在T2DM大血管和微血管并发症演变中的重要作用。
背景
就健康影响而言,最常见的血管疾病是2型糖尿病(T2DM)相关的微血管病和大血管病。这是美国死亡和发病的主要原因之一,近50%的人死于某种形式的心血管疾病。糖尿病的血管并发症同样是导致死亡的首要原因,随着血管病变相关研究不断增加,对血管疾病过程发展的原因、影响和潜在机制的全面理解是当前的主要需求。最近研究证明了表观遗传机制(如DNA甲基化和长期高血糖引起的组蛋白修饰)在糖尿病组织中基因和环境间复杂互作中的关键作用。启动子DNA甲基化诱导的基因沉默是糖尿病血管病中的最常见表观遗传修饰。这种高血糖诱导的基因启动子高甲基化有助于“代谢记忆”,即使在实现血糖控制后也会导致糖尿病的血管功能失调。另外非编码RNA或MicroRNA(miRNA)也是表观遗传学的一部分。本综述将讨论与糖尿病血管病变发病机制相关的表观遗传机制。
血管生理学和病理学
血管系统是由连接心脏和全身其他器官和组织血管组成的器官系统。血管按主题分为动脉、静脉和毛细血管三部分,动脉向组织供应来自心脏的含氧血液,静脉将未加氧的血液返回心脏进行肺氧合,毛细血管包括管理淋巴液和细胞外液的淋巴管和毛细血管。
血管病理学称为血管病变,在几乎所有血管病变病例中,从血管炎到动脉粥样硬化,内皮层的作用都密切相关。
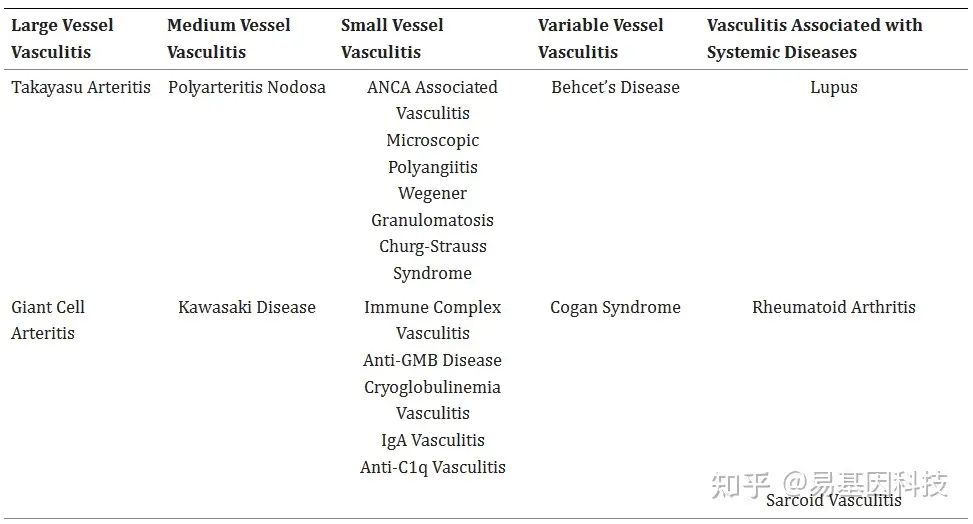
表1:基于血管大小和症状表现组织的血管炎
糖尿病和血管病变
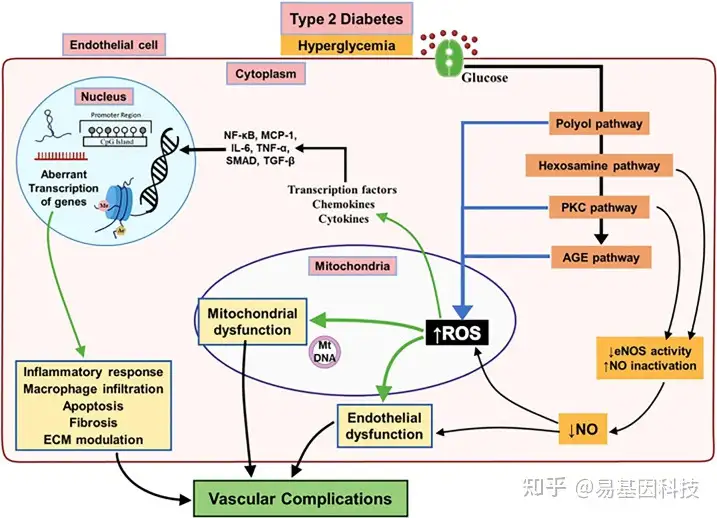
图1:导致与T2DM相关血管并发症的内皮功能失调的主要生化和细胞通路。
慢性高血糖症激活T2DM受试者内皮功能失调的不同通路,导致eNOS活性降低和线粒体ROS过量产生增加。通过激活几种改变内皮表观基因组的介质以产生促炎环境,随后调控炎症反应、巨噬细胞浸润、细胞凋亡和纤维化基因异常转录,从而导致糖尿病血管并发症。
表观遗传学和糖尿病血管病变
表观遗传修饰指不是由于DNA序列变化引起的基因转录变化。这些修饰主要调控DNA周围的核小体排列并调控基因激活或失活。本文将讨论的五种表观遗传修饰:① DNA甲基化、② 组蛋白修饰、③ 染色质重塑、④ microRNA、⑤ 长链非编码RNA(图2)。这五种不同的情况表明DNA序列域之外的外部因素可以调控复杂的疾病过程。2型糖尿病(T2DM)具有非常复杂的遗传模式,遗传和环境因素之间存在复杂的交叉。这种相互作用模拟了表观遗传修饰的功能,并且在糖尿病和糖尿病血管病变的发展中具有强烈的表观遗传作用(图3)。
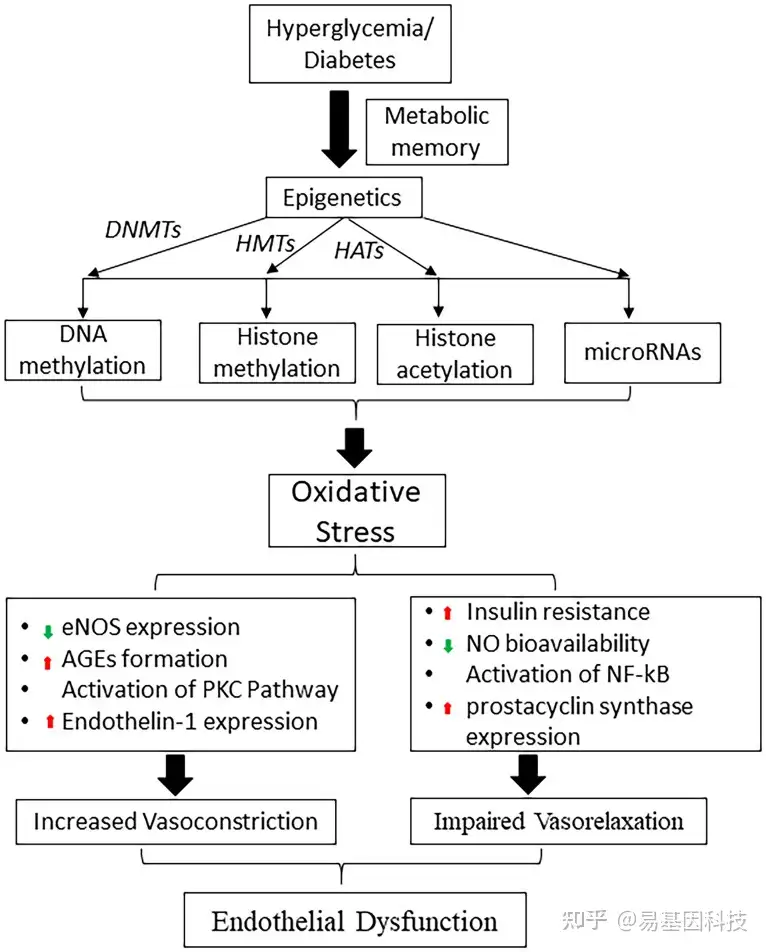
图2:高血糖诱导的表观遗传机制在代谢记忆和糖尿病内皮功能失调中的作用示意图。
DNMT:DNA甲基转移酶;HMT:组蛋白甲基转移酶;HAT:组蛋白乙酰转移酶;eNOS:内皮一氧化氮合酶;NO:一氧化氮;AGE:晚期糖基化终末产物(advanced glycation end products);PKC:蛋白激酶;NF-κB:核因子-κB。
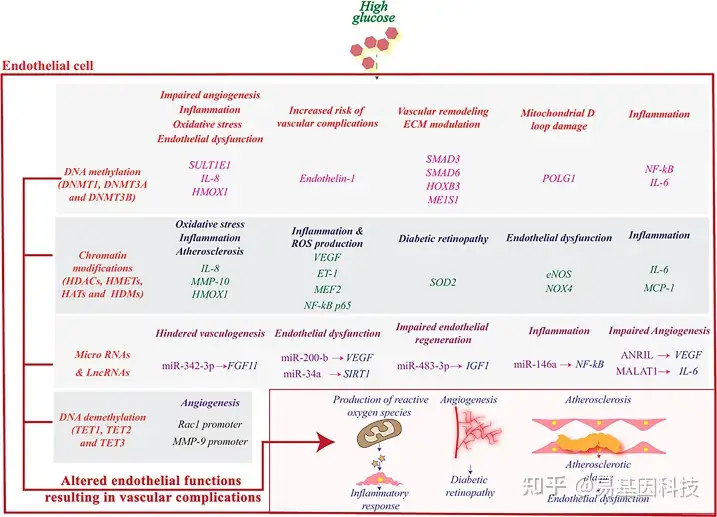
图3:高血糖诱导内皮细胞的表观遗传重编程,并影响与其功能失调相关的基因。
DNA甲基化和去甲基化、组蛋白修饰、miRNA和lncRNA调控与血管生成相关的各种基因的活性,并刺激病理生理通路、诱导炎症反应、随后导致血管并发症。
糖尿病血管病变的表观遗传机制
关于糖尿病血管病变中发生的特定表观遗传修饰以及它如何适应微/大血管病变,目前与糖尿病血管病变相关的研究文献非常多。Chen等人在2016年进行的一项大型全方位研究探索了血糖控制不佳在糖尿病微血管并发症中的作用,研究人员利用DCCT试验(糖尿病控制和并发症试验(1983年-1993年))表明严格血糖控制组和对照组之间的微血管变化非常显著。没有血糖调控的对照组显示出明显的血管病并发症(视网膜病变、肾病)迹象增加,且对照组和血糖控制组的DNA甲基化位点不同。这项研究为表观遗传学在糖尿病中的作用奠定了基础,并证实了先前的研究,即不受控制的高血糖与血管紊乱中的作用相关。
Nakatochi和研究人员发现,在心肌梗塞(糖尿病共病率超40%)中的3个特异性位点具有DNA甲基化特征。日本男性中更为相关的分析表明这些位点的DNA甲基化不仅可以预测心肌梗死,还可以预测BMI和血脂水平,这表明可以通过表观遗传学独立调控大血管疾病。Bell等人在192名爱尔兰糖尿病患者中发现19个CpG位点与糖尿病肾病相关。在Nakatochi等人报道的大血管水平和Bell等人报道的微血管水平上的表观遗传调控机制在不同人口群体中都存在,并与年龄和合并症相匹配。
糖尿病视网膜病变属于糖尿病的微血管并发症,Berdasco等研究人员检测了视网膜血管的渗出性和缺血性损伤,这两种变化都与糖尿病视网膜病变的慢性问题以及通过促血管生成因子在糖尿病控制不佳后进展失明相一致。在一个三步模型中,研究人员发现与渗出性和缺血性损伤一致的增殖性视网膜病变中有46个CpG岛甲基化标记的基因,且在启动子附近发现了整个MAP3K1通路中低甲基化。调控基因的低甲基化与基因表达上调一致,表明该通路的基因表达上调与增殖性视网膜病变增加相关。Argadh等人绘制了与该疾病相关的全面DNA基因组图谱,检测了超300个CpG位点的DNA甲基化率,这一发现不仅与糖尿病视网膜病变患者甲基化率的增加一致,而且甲基化率甚至可以作为视网膜病变严重程度的预测算法。
目前对组蛋白修饰和染色质重塑在糖尿病和血管疾病发病机制中作用的研究较为有限。在高血糖条件下,胰岛特异性转录因子Pdx1招募共激活因子p300和组蛋白甲基转移酶(HMT)SET7/9,以增加组蛋白乙酰化和H3K4me2,从而导致在胰岛素启动子处形成开放染色质并刺激胰岛素产生。另一方面在低血糖条件下,相同的转录因子Pdx1可以招募共抑制因子HDAC1/2,导致胰岛素基因表达的抑制。糖尿病组织中的持续性炎症通过增加组蛋白赖氨酸乙酰化以上调内皮细胞中的炎症基因表达,如H3K9/14Ac和组蛋白乙酰转移酶(HATs)CBP/p300在糖尿病组织的炎症基因表达中起关键作用。总之这些发现表明糖尿病刺激可以触发启动子甲基化和染色质结构变化,从而对靶基因表达产生长期影响。
糖尿病患者创面难以愈合的表观遗传学调控机制
除了血管病变外,2型糖尿病(T2DM)还有另一个主要慢性并发症:血管病变引起的溃疡。虽然不是直接的血管损伤,但难以愈合的糖尿病溃疡是与动脉供血不足和灌注减少相关性缺血的直接结果。特别是由于糖尿病氧化应激和异常的基因-环境互作,伤口愈合的皮肤机制不平衡。如前所述,内皮细胞功能失调和糖基化产物积累对瘢痕形成和愈合的逐步过程造成损害,在皮肤伤口愈合过程中观察到多种表观遗传机制,尤其以 DNA甲基化和组蛋白修饰最为显著。如在小鼠皮肤伤口中观察到H3K27me3减少,这与H3K27特异性赖氨酸去甲基化酶Jmjd3和Utx的表达增加有关;另外多梳抑制复合物2(PRC2)的组分:Eed、Ezh2和Suz12(其甲基化H3K27)在小鼠伤口愈合期间被下调;还有一个严格受表观遗传调控的基因是eNOS,在生理条件下内皮细胞通过沿对称链和CpG二核苷酸的大部分低甲基化启动子区域显示eNOS的持续激活。Yan等人在2010年的研究表明内皮细胞和非内皮细胞的eNOS染色质结构也不同,且组蛋白去乙酰化酶(HDAC)抑制剂可以上调eNOS水平。因此在糖尿病伤口中发现的eNOS水平失调显示了表观遗传调控在伤口愈合中的作用的直接通路。
由于在未愈合的伤口中发现的功能失调直接与单核细胞趋化蛋白-1(MCP-1)的基因水平有关,因此巨噬细胞的作用也受到质疑。与对照小鼠相比,在链脲佐菌素(streptozotocin)诱导的糖尿病巨噬细胞中的MCP-1基因上调,上调的表达通过MCP-1启动子区单甲基化直接受表观遗传调控。染色质重塑是参与调控炎症相关基因转录的重要表观遗传修饰之一,炎症相关基因影响巨噬细胞极化和其他对伤口愈合至关重要的特性。具体而言,HAT和组蛋白去乙酰化酶(HDAC)激活炎性单核细胞分化和巨噬细胞表型,组蛋白修饰酶对巨噬细胞相关基因的改变与伤口愈合受损相关;ATP依赖性染色质重塑(SWI-SNF)复合体在巨噬细胞发育中起作用;SWI-SNF与HDAC1互作以调控组蛋白乙酰转移酶(H3K27ac)并调控对细胞发育和分化至关重要的基因。
基于非编码RNA的治疗在糖尿病伤口愈合中应用
MicroRNA(miRNA)是短的单链,与影响基因表达的mRNA强烈结合。miRNA的治疗潜力值得探索:
- 单个miRNA可以通过调控多个功能趋同的靶基因而起到放大器的作用;
- miRNA是可以用新技术操作的稳定的小生物分子;
- 可以以受控的方式精确传递,治疗效果可以通过过度表达特定miRNA或使其沉默来实现。
通过使用基于纳米技术的新兴方法,使用病毒或非病毒方法进行基因治疗,可以实现潜在治疗性生物分子传递。研究表明miRNA功能失调与糖尿病发病机制和并发症相关(图4)。如内皮细胞已被证明在严重高血糖症中具有特异性上调的miRNA,而在糖尿病足溃疡的血浆中具有类似的上调。Madhyastha等人的研究发现,与对照小鼠相比,14种miRNA在糖尿病条件下具有表达变化,特别是在糖尿病伤口愈合期间,miR-146b上调了近30倍。而MiR-21在糖尿病伤口愈合期间下调。Li等人在2009年发现,利用miR-221抑制剂治疗使miR-221水平下调,并在高血糖条件下改善了细胞迁移。
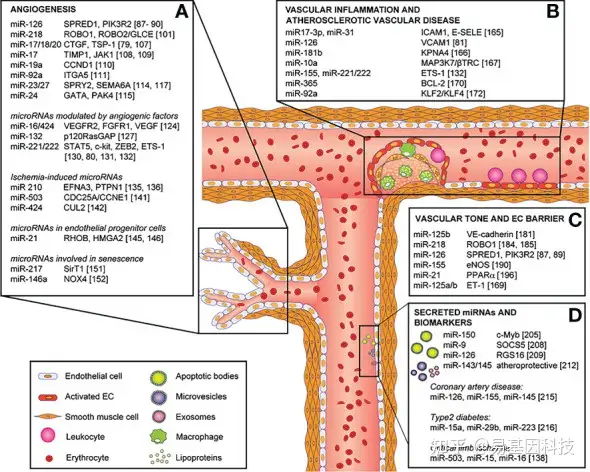
图4:miRNA在内皮细胞表型、功能和血管疾病及其直接靶点中的作用。
miRNA包括:(A)血管生成,(B)血管炎症和动脉粥样硬化性血管疾病,(C)血管张力和内皮细胞屏障,(D)分泌的miRNA和生物标志物。
MiR-200b长期以来一直与糖尿病血管病变有关。先前研究表明miR-200b在糖尿病伤口的血管生成中具有调节作用。然而miRNA在高血糖中的调节作用存在问题。本综述通过表观遗传修饰探讨了高血糖对miR-200b启动子区的作用。结果发现在严重高血糖的糖尿病伤口内皮细胞中,miR-200b启动子区域存在明显的低甲基化,这表明了miR-200b低甲基化状态的第一个证据,且这一事实被甲基供体S-腺苷-L-蛋氨酸通过重新甲基化启动子区域来回复内皮功能所证实。因此,调控特定miRNA表达可能是克服糖尿病相关并发症的有效治疗方法。特定基因的过表达和下调主要由miRNA调节的创伤生物学调控,解开皮肤伤口愈合中miRNA失调过程将有助于开发新的靶向疗法。OxymiRs是响应组织氧合状态的miRNA,目前已经研究了在伤口愈合过程中差异表达的多种oxymiR。缺氧敏感的miRNA称为“HypoxymiRs”,慢性非愈合伤口(如糖尿病足溃疡、静脉溃疡和压疮)以缺血/缺氧为特征。miR-203、miR-210和miR-21是被充分研究的积极参与伤口愈合的“HypoxymiRs”。同时,miRNA在伤口愈合过程中的炎症调控中起着非常关键的作用。miR-155、miR-146a和miR-132是一些与伤口和炎症相关的miRNA,miR-155调节参与针对病原体的免疫反应的蛋白质表达,这在慢性感染伤口中具有临床意义,它还通过其他信号介质参与调节TNFα。miR-125b、miR-31、miR-17-3p和miR-124a是创伤愈合中参与炎症调节的其他miRNA。miRNA在皮肤伤口的上皮化过程中的维持屏障功能方面发挥关键作用,miR-210依赖性通路损害缺血伤口上皮化。皮肤角质形成细胞中miR-1过表达会损害细胞迁移,从而影响上皮化和皮肤屏障功能。miRNA在伤口愈合过程中通过诱导血管形成在伤口愈合期间的血管生成反应中起着重要作用。多种miRNA可以参与不同阶段的血管生成过程:内皮细胞的增殖、迁移和形态发生。参与血管生成的miRNA也称为血管miR,包括miR-15b、miR-16、miR-20、miR-21、miR-23a和其他,这些miRNA可通过基因治疗方法调控其表达而应用于临床。然而现有的挑战是以活性形式传递至相关组织/器官的有效性和特异性。在理想的条件下,所传递的miRNA细胞摄取应该很高且没有内体逃逸。另外靶向miRNA的治疗方法可能由于对多个基因的下游作用而产生意外脱靶效应。
长链非编码RNA(LncRNAs)是另一组在血管信号传导中发挥重要作用的非编码RNA。LncRNA通过调节染色质动力学和转录活性来调控基因表达。LncRNA在心血管疾病、糖尿病和原发性开角型青光眼等疾病中明显调控失败。如lncRNA ZEB-AS1充当miR-200b海绵可以调节细胞迁移、侵袭和增殖。但这些lncRNA也可能导致T2DM疾病或其他相关糖尿病相关并发症进展。越来越多证据表明多个lncRNA与糖尿病并发症有关,多个血管生成miRNA‐lncRNA对与成熟期的伤口愈合有关。在伤口愈合过程中, LncRNAs GAS5、IGF2AS、MALAT1、ANRIL、H19、MIAT和lncEGFL7OS调节血管生成过程。此外 T2DM患者的lncRNA NKILA、NEAT1、MALAT和MIAT表达增加可能影响T2DM患者的疾病程度和严重程度。因此,糖尿病伤口血管生成在严格调节的表观遗传调控下进行。
结论
血管病理学的范围包括急性疾病和慢性衰弱疾病。T2DM和糖尿病血管病围绕复杂的化学级联和基因组发现揭示了遗传学在疾病过程中的作用。从大血管动脉粥样硬化到小的外周内膜增生和内皮功能失调,糖尿病血管病涉及多器官病理,其治疗干预有限。但目前仍缺乏对糖尿病血管病变调控机制类型的全面综述。具体而言,DNA中外在发生的糖尿病调控过程尚未得到充分研究。其中表观遗传学对糖尿病的研究非常有意义。本文综述了糖尿病血管病的表观遗传调控,这些调控过程完全发生在DNA结构和序列之外,因此对其潜在的治疗标靶很有吸引力。
对糖尿病血管病变的表观遗传调控研究表明相关的表观调控因子具有多种作用。血管生成丢失不仅影响糖尿病伤口,还可能导致灌注受限、全身性高血压和皮肤伤口愈合,而糖尿病视网膜病变中血管生成的增加则与血管增生导致视网膜病变恶化的主题相反。因此当表观遗传修饰在体内发生变化时,仍需要阐明其具体作用。microRNA具有强大的调节功能,但其功能在很大程度上取决于高血糖等条件。表观遗传学的作用本身就是药物干预的主要疗法。由于这些机制在基因组之外运行且没有下游级联,因此这些甲基化蛋白或组蛋白修饰变化可以对糖尿病血管病变的治疗产生深远影响。
易基因科技提供全面的表观组学测序服务,有DNA甲基化、组蛋白修饰、RNA甲基化测序需要的老师可联系。

参考文献:Bhamidipati T, Kumar M, Verma SS, Mohanty SK, Kacar S, Reese D, Martinez MM, Kamocka MM, Dunn KW, Sen CK, Singh K. Epigenetic basis of diabetic vasculopathy. Front Endocrinol (Lausanne). 2022;13:989844.
相关文献:

 浙公网安备 33010602011771号
浙公网安备 33010602011771号